翼龍
(翼龍,
,
그리스어
πτερ?σαυρο?,
pterosauros,
프테로社友로스에서 온 말로 "날개달린 도마뱀"이라는 意味)은 "翼龍" 盆地群, 或은 "翼龍"
목
에 該當하는 하늘을 나는 爬蟲類이다.
[1]
[2]
[3]
翼龍은 大衆媒體와 一般的인 사람들에 依해 흔히 하늘을 나는
恐龍
으로 指稱되곤 하지만
支配爬蟲類
에 屬하는 하늘을 나는 恐龍이 아닌
動物
들을 가리키는 말이다. 現代의 科學界의 意見은
魚龍
이나
수장룡
을 비롯한 다양한 種類의 滅種한 海洋
爬蟲類
들은 勿論 翼龍 亦是 여기(恐龍)에 包含되지 않는다는 것이다.
[4]
特徵
[
編輯
]
翼龍은 解剖學的으로 볼 때 飛行을 위해 祖上인 爬蟲類로부터 매우 큰 變化를 겪었다. 翼龍의
骨格
은
새
의 뼈와 마찬가지로 속이 비었고 空氣로 차 있다.
가슴뼈
에는 飛行을 위한
筋肉
이 附着될 수 있도록
龍骨突起
가 發達해 있으며 커다란
腦
는 飛行과 關聯하여 特化된 機能을 가지고 있었던 것으로 보인다.
[5]
後期의 翼龍에서는 어깨 위의 脊椎뼈가 합쳐져 '노타리움'(notarium)이라고 불리는 뼈가 되어 飛行 中에 몸통을 단단하게 해주고
肩胛骨
을 安定的으로 支持해 주었다.
날개
[
編輯
]
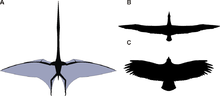 퀘찰코아틀루스
노르트롭아이의 날개 復元도(A)를
信天翁
디오메데아 엑술란스(
Diomedea exulans
)(B), 그리고 안데스
콘도르
(
Vultur gryphys
)와 比較한 그림. (크기는 正確하지 않음)
퀘찰코아틀루스
노르트롭아이의 날개 復元도(A)를
信天翁
디오메데아 엑술란스(
Diomedea exulans
)(B), 그리고 안데스
콘도르
(
Vultur gryphys
)와 比較한 그림. (크기는 正確하지 않음)
翼龍의 날개는 皮膚와 其他 組織들로 이루어진 飛膜으로 되어 있다. 飛膜은 各 팔에서 가장 긴 네番째
손가락
에서부터 몸의 兩 옆을 따라 발목까지 連結되어 있다.
歷史的으로 翼龍의 날개는 皮膚로 이루어진 가죽같은 構造로 매우 單純한 것이라고 생각되어 왔으나 硏究에 依해 비막이 實際로는 매우 複雜하고 力動的인 構造로 能動的인 形態의 飛行에 적합한 것임이 밝혀졌다. 바깥쪽 날개(날개의 끝部分에서 팔꿈치까지)는 '額티盧피브릴'이라고 불리는 纖維가 촘촘히 자리잡아 날개를 强化해 준다.
[6]
額티盧피브릴 自體는 세 個의 뚜렷히 區分되는 層이 서로 다른 方向으로 겹쳐져 있어 十字模樣 패턴을 形成한다. 額티盧피브릴의 實際 機能은 勿論 이것이 正確히 어떤 物質로 만들어져 있었는지도 알려져 있지 않다. 正確한 構成 (케라틴, 筋肉, 彈力性 構造 等等) 에 따라 날개의 바깥쪽 部分을 단단하게, 或은 탄탄하게 만들어주는 役割을 했을 것이다.
[7]
飛膜은 또 얇은 근육층과 纖維質 組織, 그리고 고리를 이루는 핏줄로 된 獨特하고 複雜한 循環系를 가지고 있다.
[8]
大型 翼龍의 날개뼈에서 보이는 속이 비어 있는 構造와 最小限 한 個의 標本에 保存된 軟質部에서 알 수 있듯이 어떤 翼龍은 呼吸에 使用되는 氣囊 (아래의 古生物學 部分을 볼 것) 시스템을 飛膜에까지 擴張시켰다.
[9]
翼龍의 날개 構成要素
[
編輯
]
 이 그림에서
소르데스
가 描寫된 것을 보면, 翼龍의 飛膜은 다리와 連結되어 있었으나 박쥐와 달리 꼬리까지는 連結되지 않았을 것으로 보인다.
이 그림에서
소르데스
가 描寫된 것을 보면, 翼龍의 飛膜은 다리와 連結되어 있었으나 박쥐와 달리 꼬리까지는 連結되지 않았을 것으로 보인다.
翼龍의 飛膜은 세 部分으로 나뉜다. 첫番째는 날개의 가장 앞部分인 "前飛膜(propatagium)"으로 손목과 어깨 사이에 位置하며 飛行 時에 全然(leading edge)을 이룬다. 이 部分은 어떤 標本에서 볼 수 있는 것처럼 첫 세 손가락을 包含할 수도 있다.
[8]
"팔飛膜(brachiopatagium)"은 날개의 주된 構成要素로 길게 늘어난 네番째 손가락과 뒷다리 (뒷다리의 正確히 어느 部分까지인지는 確實하지 않으며 種에 따라 달랐을 수 있다. 아래를 볼 것) 사이를 連結한다. 마지막으로 最小限 一部 翼龍 그룹들은 다리 사이, 어쩌면 꼬리까지 連結하는 "꼬리飛膜(uropatagium)"을 가지고 있었다. 꼬리飛膜이 어디까지 連結되어 있었는지는 분명하지 않은데,
소르데스
에 對한 硏究에서는 다리 사이를 連結하지만 꼬리는 包含되지 않는 것으로(이 境遇 "다리飛膜(cruropatagium)"이라고 할 수 있음) 보았다
프테로닥틸루스類가 아닌 翼龍
은 넓은 꼬리/다리飛膜을 가지고 있었으나 프테로닥틸루스類는 다리 사이에만 飛膜을 가지고 있었다는 데 多數의 學者들이 同意하고 있다.
프테라노돈
은 特히 꼬리의 形態로 볼 때 꼬리飛膜을 가지고 있었던 것이 確實해 보인다.
 날개의 構造
날개의 構造
翼型骨(pteroid)로 알려진 뼈는 翼龍에게만 있는 것으로 손목에 連結되어 손목과 어깨 사이에서 前飛膜을 받쳐준다. 翼龍의 첫 세 個 손가락 사이에 膜이 있었던 痕跡이 있는데, 이것은 前飛膜이 예전의 復元圖에서 흔히 그려졌던 것처럼 익형골에서 어깨까지 이어지는 것이 아니라 더 넓은 範圍에 걸쳐 있었으리라는 것을 示唆한다.
[8]
翼型골의 位置 自體에 對해서도 論爭이 있었다. 매튜 윌킨슨과 같은 科學者들은 翼型골이 앞쪽으로 뻗어 있어 前飛膜을 길게 늘렸을 것이라고 主張했다.
[10]
하지만 이 觀點은 크리스 베넷의 2007年 論文에서 强力한 批判을 받았다. 크리스 베넷은 翼型골이 以前에 생각했던 것처럼 다른 뼈와 連結되지 않으며 앞으로 뻗어 있을 수 없다는 것을 보였고, 傳統的으로 생각되어왔던 것처럼 몸 쪽을 向했다는 것을 보였다.
[11]
形態學的, 發生學的, 그리고 組織學的 證據를 살펴보면 翼型골은 骨質化된 軟骨이 아니라 眞짜 뼈였다. 翼型골의 起源은 분명하지 않다. 손목뼈가 變形된 것일 수도 있고, 손바닥뼈, 或은 새로 만들어진 뼈日 수도 있다.
[12]
翼龍의 손목은 翼型골을 除外하면 두 個의 몸쪽 손목뼈와 네 個의 바깥쪽 손목뼈로 이루어져 있다. 聖體 標本에서는 몸쪽 손목뼈가 하나로 합쳐져 "新카팔(syncarpal)"을 이루며 바깥쪽 손목뼈 中 세 個가 합쳐져 바깥쪽 新카팔을 이룬다. 하나 남은 바깥쪽 손목뼈는 中央손목뼈 (medial carpal)로 불리며 바깥쪽 新카팔의 앞쪽 表面에 位置한 긴 隆起部와 連結된다. 中央손목뼈에는 움푹 들어간 部分이 앞쪽, 아래쪽, 그리고 몸 中央 쪽에 있는 데 이곳에 翼型골이 連結된다.
[13]
古生物學者들 사이에서 翼龍의 週 飛膜人 팔飛膜이 다리에 連結되는지, 連結된다면 어느 部分에 連結되는지에 對해서는 相當한 論爭이 있어 왔다. 람포린쿠스類인
소르데스
[14]
,
아누로그나티드
人
제홀옵테루스
,
[15]
, 그리고
산타나 層
에서 發見된 프테로닥틸루스類를 보면 最小限 一部 種에서는 비막이 다리까지 連結되지 않았던 것으로 보인다.
[16]
하지만 現生
박쥐
와
날다람쥐
를 보면 飛膜의 範圍는 相當한 變異가 있기 때문에 이들처럼 서로 다른 種類의 翼龍들이 서로 다른 날개 模樣을 가지고 있었을 수 있다. 實際로 翼龍 팔다리의 比率을 보면 相當한 變異가 있으며 이것은 날개의 構造가 다양했다는 意味일 수 있다.
[17]
全部는 아니었을 수 있지만 많은 種類의 翼龍은 발에 물갈퀴를 가지고 있었다.
[18]
頭蓋骨, 이빨 그리고 볏
[
編輯
]
 아마도
콜로保隣쿠스
(
Coloborhynchus
)의 것으로 보이는 이빨
아마도
콜로保隣쿠스
(
Coloborhynchus
)의 것으로 보이는 이빨
거의 大部分의 翼龍 頭蓋骨은 긴 턱과 바늘같은 이빨을 가지고 있다. 어떤 境遇에는 化石化된
케라틴
質 부리가 保存되어 있기도 하지만 이빨을 가진 翼龍의 境遇 부리는 턱의 끝部分에 작게 位置하고 있으며 이빨은 부리에 包含되지 않는다.
[19]
부리를 가진 形態 中 發展된 種類는
프테라노돈과
와
아즈다르코類과
의 境遇처럼 이빨이 없으며 더 크고 새와 類似한 부리를 가지고 있다.
[20]
頭蓋骨의 눈 앞쪽에 여러 個의 구멍을 가진 거의 大部分의
支配爬蟲類
와는 달리 프테로닥틸루스類 翼龍은
奠雁蝸瘡
과 콧구멍이 하나의 커다란 구멍으로 합쳐저 "前眼窩悲愴"(nasoantorbital fenestra) 이라고 불린다. 하늘을 날아야 하기 때문에 가벼운 頭蓋骨을 만들기 위해 이런 特徵이 鎭火했을 수 있다.
[20]
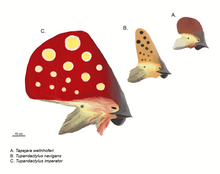 볏의 復元도: 볏을 가지고 있는
타페야라과
翼龍 세 種類. 위에서부터 次例로
타페야라
웰른호퍼아이(
Tapejara wellnhoferi
), "타페야라" 나비간스("Tapejara"
navigans
),
투판닥틸루스 임페라토르
(
Tupandactylus imperator
)
볏의 復元도: 볏을 가지고 있는
타페야라과
翼龍 세 種類. 위에서부터 次例로
타페야라
웰른호퍼아이(
Tapejara wellnhoferi
), "타페야라" 나비간스("Tapejara"
navigans
),
투판닥틸루스 임페라토르
(
Tupandactylus imperator
)
翼龍은 華麗한 볏을 가지고 있는 境遇가 많다는 것이 잘 알려져 있다. 第一 처음, 그리고 아마도 가장 잘 알려진 例는
프테라노돈
종들의 뒤로 뻗어나온 볏이지만, 몇몇
他페하리드과
翼龍과
닉土社友루스
等 몇몇 種類는 뼈로 만들어진 볏의 基底部에 더해 케라틴이나 다른 軟質部로 만들어진 宏壯히 커다란 볏을 가지고 있었을 것이다.
1990年代부터 새로 發見된 翼龍은 勿論 오래된 標本들을 더 仔細히 調査하면서 볏이 翼龍들 사이에서 예전에 생각했던 것보다 훨씬 더 널리 퍼진 特徵이었다는 것이 알려졌다. 많은 境遇 볏이 케라틴으로 延長되어 있거나, 때로는 케라틴만으로만 이루어진 볏이 있어서 化石化되지 않는 境遇가 있기 때문이다.
[8]
프테로린쿠스
(
Pterorhynchus
)나
프테로닥틸루스
(
Pterodactylus
)의 境遇와 같이 볏이 實際로 어느 程度 크기였는지는
紫外線
寫眞을 통해서만 알 수 있다.
[19]
[21]
프테로린쿠스와
아우스트리아닥틸루스
(
Austriadactylus
)는 모두 볏을 가진
람포린쿠스
流로, 原始的인 翼龍조차도 볏을 가지고 있다는 것을 보여주고 있다. (예전에는 더 發展된 形態인 프테로닥틸루스類 翼龍만 볏을 가지고 있다고 생각했다)
[8]
피크老파이버
[
編輯
]
最小限 몇몇 翼龍들은 '피크老파이버'로 알려진 가는 실 模樣의
털
같은 것을 머리와 몸에 가지고 있었다.
哺乳類
의 털과는 類似하지만
상동
(homologous), 卽 共通의 起源을 가지는 構造物은 아니었을 것이다. 털과 비슷하게 보송보송한 外皮(integument)를 가지고 있었다는 것은 1831年에 골드퍼스에 依해 처음 報告되었고
[22]
, 最近에 發見된 標本들과
組織學
技術의 發展 및 標本에 對한 紫外線 檢査에 힘입어 確實한 證據를 가지게 되었다. 卽, 翼龍은 피크老파이버로 된 外皮를 가지고 있었다. 피크老파이버는 哺乳類에서 볼 수 있는 것과 같은 種類의 털은 아니지만, 겉보기에 類似한 形態로 發達한 獨特한 構造다. 어떤 境遇에는 飛膜의 額티盧피브릴 (內部의 纖維質 救助)李 피크老파이버라 眞짜 털과 混同되는 境遇가 있으나
소르데스
(
Sordes pilosus
, 털이 많은 惡魔라는 意味)나
제홀옵테루스
(
Jeholopterus ninchengensis
)의 化石 같은 境遇는 理論의 餘地가 없는 피크老파이버 자국이 머리와 몸 部分에 남아 있다. 박쥐와도 類似한데,
收斂進化
의 한 例라고 할 수 있다.
[14]
머리 部分의 피크老파이버는 只今까지 發見된 標本들 中 많운 수가 가지고 있는 커다란 턱까지는 덮고 있지 않다.
[22]
어떤 硏究者들은 (제르카스와 誌의 2002年 論文 같은 境遇) 피크老파이버가 遠視깃털의 先行形態가 아닌가 생각하기도 하지만 現在 알려져 있는 翼龍의 外皮는 새를 비롯한
마니랍토라
恐龍의 化石 記錄에서 發見되는 깃털과는 確實히 다르다.
[22]
翼龍의 피크老파이버는 遠視깃털과는 다른 構造를 가지고 있다.
[7]
[23]
피크老파이버는 柔軟하고 짧은 纖維質로 "어떤 標本에서는 5-7mm 程度에 不過下"며, 比較的 單純하여 "中央의 管(canal)을 除外하면 그 어떤 內部構造도 存在하지 않는 것으로 보인다."
[22]
翼龍의 "털가죽"은 "化石化된 哺乳類에서 發見된 것과 類似하게 纖維로 촘촘하게 덮인 狀態로 保存되어" 있었던 것으로 보아, 다른
中生代
哺乳類의 털가죽과 比肩될 만한 두께로 피크老파이버가 翼龍의 몸을 덮고 있었으리라 推定된다. 털의 두께와 털로 덮여 있던 表面의 넓이는 翼龍 種類에 따라 달랐을 것이고, 날개에서는 피크老파이버가 發見된 적이 없다.
피크老파이버가 있다는 것, 그리고 飛行을 했다는 것은 翼龍이
耐熱性
(溫血)이었을 可能性이 높다는 것을 暗示한다. 翼龍 날개에 피크老파이버가 없는 것으로 보다 털에
氣體力學
的인 機能은 없었을 것이며
耐熱性
新陳代謝에 依해 發生된 熱을 保存하는데 斷熱이 必須的이기 때문에 피크老파이버가 翼龍의 體溫調節을 爲한 것이라는 아이디어를 뒷받침해준다.
[22]
翼龍의 "털"은 매우 獨特해서 哺乳類의 털가죽이나 다른 動物의 外皮와 뚜렷하게 區分되기 때문에 새로운 이름이 必要했다. "피크老파이버"는 "촘촘한 纖維"라는 意味로 제홀옵테루스의 軟質部 자국을 다룬 古生物學者 알렉산더 켈너(Alexander W. A. Kellner)와 同僚들의 2009年 論文에서 처음 만들어진 單語다.
[7]
發見의 歷史
[
編輯
]
 프테로닥틸루스
안티쿠스(
Pterodactylus antiquus
)의 완모式標本 版畫. 1784年 에기드 베르헬스트 2 世(Egid Verhelst II) 作品
프테로닥틸루스
안티쿠스(
Pterodactylus antiquus
)의 완모式標本 版畫. 1784年 에기드 베르헬스트 2 世(Egid Verhelst II) 作品
最初의 翼龍
化石
은 1784年
이탈리아人
博物學者인
코시모 콜리니
에 依해 記載되었다. 콜리니는 翼龍 標本을 바다에 살던 動物로 긴 앞다리를 盧처럼 使用했던 것으로 잘못 解釋했다.
[24]
몇몇 科學者들은 1830年에 獨逸 動物學者
요한 게오르그 바글러
가 프테로닥틸루스의 날개가 물갈퀴처럼 利用되었을 것이라는 提案을 할 때까지 이 解釋을 支持했다.
[25]
1801年에
조르쥬 퀴비에
가 처음으로 翼龍이 하늘을 날던 動物이라고 提案했으며,
[26]
1809年에는 獨逸에서 發見된 標本에 "프테로-닥틸"이라는 이름을 붙였다.
[27]
學名이 標準化되면서 이 속의 이름은
프테로닥틸루스
(
Pterodactylus
)가 되었지만 "프테로닥틸"이라는 이름은 大衆的으로 繼續 쓰였고 翼龍 全體를 가리키는 名稱으로도 不正確하게 쓰이고 있다.
[28]
古生物學者들은 "프테로닥틸"이라는 單語를 使用하는 代身 "翼龍(pterosaur)"이라는 用語를 選好한다. "프테로닥틸"이라는 用語는 "프테로닥틸루스" 속에 屬하는 翼龍들, 或은 좀 더 넓은 意味로 프테로닥틸루스亞目의 翼龍을 가리키는 것으로 使用된다.
[29]
 三次元的으로 保存된 브라질 산타나 層의
안항구에라
(
Anhanguera santanae
)의 頭蓋骨.
三次元的으로 保存된 브라질 산타나 層의
안항구에라
(
Anhanguera santanae
)의 頭蓋骨.
古生物學
[
編輯
]
飛行
[
編輯
]
 四足步行에서 離陸을 하는 프테라노돈 弄氣켑스(
Pteranodon longiceps
)의 骨格 復元도
四足步行에서 離陸을 하는 프테라노돈 弄氣켑스(
Pteranodon longiceps
)의 骨格 復元도
翼龍의 飛行 技法은 아직 完全히는 理解되지 않고 있다.
[30]
[31]
 呼吸 動作(위쪽)과 內部 氣囊 시스템(아래쪽)을 보여주는 그림
呼吸 動作(위쪽)과 內部 氣囊 시스템(아래쪽)을 보여주는 그림
日本人 科學者인 사토 가쓰後尾(佐藤克文)는 現生 潮流를 利用하여 計算을 해 보고 翼龍이 空中에 떠 있는 것은 不可能하다고 結論내렸다.
[32]
"翼龍의 姿勢, 移動, 그리고 古生態"라는 冊에서는 翼龍이 하늘을 날 수 있었던 것이 酸素濃度가 높고 더 密度가 높았던
白堊紀
後期의 大氣 때문이었을 것이라는 理論을 提起했다.
[33]
하지만 가쓰後尾 및 "翼龍의 姿勢, 移動, 그리고 古生態"의 著者들은 翼龍에 對한 오래된 理論을 基盤으로 硏究를 遂行했다. 이 오래된 理論에 따르면 翼龍은 바다새와 類似하며 크기 一部 制限은 아즈다르코類과 및 他페하리드과 等 陸上生活을 하는 翼龍에는 適用되지 않는다. 거기에 더해서 대런 네이쉬는 現在와 中生代의 大氣 造成 差異가 翼龍의 巨大한 크기를 說明하기 위해 꼭 必要한 것은 아니라고 結論 내렸다.
[34]
또 한 가지 理解하기 힘든 點은 이들이 어떻게
離陸
을 했는가이다. 初期의 理論은 翼龍은 大槪 冷血性으로 칼로리를 태워 熱을 얻는 代身 現生 도마뱀처럼 周圍環境으로부터 溫氣를 얻으며 滑空을 하는 動物로 보았다. 巨大한 翼龍이 非效率的인 冷血性 新陳代謝를 하면서 뒷다리만 使用해 空中으로 뜰 수 있는 推進力을 얻는 새와 類似한 離陸 動作을 할 수 있었겠는가? 翼龍은 溫血性이었으며 强力한 飛行 筋肉을 가지고 있었고, 飛行筋肉을 利用해
네 다리로 걸었다
고 한다.
[35]
포츠머스 大學
의 마크 위튼과
존스 홉킨스 大學
의 마이크 下빕은 翼龍이 跳躍하는 方式으로 날아올랐을 것이라고 한다.
[36]
날개가 있는 앞다리는 매우 强力해서 離陸을 쉽게 해주었을 것이다.
[35]
一旦 空中에 뜨면 翼龍은 最高 時速 120 킬로미터에 達하는 速度로 數千 킬로미터씩 移動할 수 있었을 것이다.
[36]
1985年에 스미소니언 協會는 航空工學自認
폴 맥크리디
에게 折半 크기의
퀘찰코아틀루스
노르트롭아이 模型을 만들어 作動시키게 했다. 이 模型은 땅에서 윈치를 利用해 離陸했다. 1986年에 이 模型을 여러 次例 날렸고 映像은 스미소니언의 아이맥스 映畫인 "온 더 윙 (On the Wing)"의 一部가 되었다. 하지만 이 模型은 解剖學的으로 正確하지 않으며 翼龍들에게는 없었던 垂直 및 水平 安定裝置를 가지고 있었다. 또 꼬리가 實際보다 길어 무게 分布를 바꿔놓았다.
氣囊과 呼吸
[
編輯
]
2009年의 硏究에서는 翼龍이 허파-氣囊 시스템과 正確하게 制御되는 骨格 呼吸 펌프를 가지고 있어 새와 類似하게 한 方向으로 空氣가 흐르는 呼吸系를 가지고 있을 可能性이 높다고 보았다. 最小限 一部 프테로닥틸루스類는 皮下氣囊 시스템을 가지고 있어 몸무게를 더 줄여주었을 것이다.
[9]
神經系
[
編輯
]
엑스레이를 利用해 翼龍의
腦
가 들어 있던 空間을 硏究해본 結果 람포린쿠스 무엔스터아이(
Rhamphorhynchus muensteri
)와 안항구에라 산타나에(
Anhanguera santanae
)는 巨大한 小葉(flocculi)을 가지고 있었던 것으로 밝혀졌다.
小葉
은 關節, 筋肉, 皮膚 및 平衡器官 等에서 오는 信號를 綜合하는 腦의 한 部分이다.
[5]
翼龍의 小葉은 全體 腦 質量의 7.5%를 차지하고 있었으며 이것은 어떤 脊椎動物보다도 높은 比率이다. 새도 다른 動物들과 比較하면 유난히 큰 小葉을 가지고 있는데 前提 腦 質量의 1~2 퍼센트만을 차지하고 있다.
[5]
小葉은 눈 筋肉을 조금씩 自動으로 움직이게 하는 神經信號를 보낸다. 이 움직임을 통해 網膜에 맺히는 像이 흔들림 없이 維持된다. 翼龍이 巨大한 小葉을 가지고 있었던 理由는 아마 커다란 날개를 가지고 있어서 處理해야 할 感覺 情報가 많았기 때문이었을 것이다.
[5]
陸上에서의 움직임
[
編輯
]
 아즈다르코類과의 翼龍이 남긴 것으로 보이는 痕跡化石 海南이크누스 우항리엔시스(
Haenamichnus uhangriensis
)
아즈다르코類과의 翼龍이 남긴 것으로 보이는 痕跡化石 海南이크누스 우항리엔시스(
Haenamichnus uhangriensis
)
翼龍의 臗骨臼는 若干 위쪽으로 向해 힜고
넙다리뼈
의 머리 部分은 살짝 안쪽을 보고 있어서 半直立 姿勢를 取했을 것으로 보인다. 翼龍은 滑空하는 도마뱀처럼 飛行中에 허벅지를 水平으로 維持할 수 있었을 것이다.
翼龍이
四足步行
을 했는지
이族譜行
을 했는지에 對해서는 相當한 論難이 있었다. 1980年代에 古生物學者 케빈 페이디언이
디모르포돈
(
Dimorphodon
)처럼 긴 뒷다리를 가진 작은 翼龍들은 하늘을 나는 것 外에 두 다리로 걷거나 甚至於
駝鳥
처럼 달리기도 했을 것이라고 主張했다.
[37]
하지만
全羅南道
海南郡
等地에서 發見된 翼龍 발자국 化石 等 많은 數의 翼龍 步行列(trackway)에서 네 個의 발가락을 가진 뒷발과 세 個의 발가락을 가진 앞발의 자국이 모두 發見되면서 翼龍이 네 다리로 걸었다는 것이 분명해 졌다.
[38]
[39]
발뒤꿈치를 들고 발가락으로 걷는 (
趾行性
digitigrade) 거의 大部分의 脊椎動物과 달리 翼龍의 발자국은
사람
이나
곰
과 類似하게 발 全體가 땅바닥과 닿은 (
척行星
plantigrade) 形態를 보여주고 있다. 아즈다르코類과의 발자국을 보면 最小限 몇몇 翼龍들은 네 다리가 몸의 아래쪽으로 뻗어 있는 直立步行을 했다는 것을 알 수 있다. 에너지 效率이 높은 直立步行은 옆으로 다리를 뻗는 現生 爬蟲類의 步行 形態와는 달리 거의 大部分의 現生 鳥類와 哺乳類가 使用하는 步行 形態이다.
[18]
[35]
 步行列 化石은
퀘찰코아틀루스
노르트롭아이 같은 翼龍들이 네 발로 걸었다는 것을 보여준다
步行列 化石은
퀘찰코아틀루스
노르트롭아이 같은 翼龍들이 네 발로 걸었다는 것을 보여준다
傳統的으로 땅 위에서의 모습은 語塞하고 우스꽝스럽게 그려졌지만 翼龍의 (特히 프테로닥틸루스類의) 解剖學的 特徵들을 보면 이들은 걷거나 뛰는 데 能熟했던 것으로 보인다.
[40]
아즈다르코類과 및 오르泥土케이리드과 翼龍의 앞다리 뼈는 다른 翼龍들과 比較해 보면 唯獨 길고, 아즈다르코類科에서는 팔의 뼈와 손 (中手骨) 이 特別히 길쭉하다. 거기에 더해 全般的으로 아즈다르코類과의 앞다리는 그 比率이 빠른 速度로 달리는
有蹄類
哺乳動物과 類似하다. 反面, 이들의 뒷다리는 빠른 速度를 낼 수 있는 構造는 아니지만 다른 翼龍들과 比較하면 길어서 步幅이 컸을 것이다. 아즈다르코類과 翼龍들은 아마도 달리지는 못했겠지만 相對的으로 빠르고 效率的인 움직임을 보였을 것이다.
[18]
翼龍 손과 발의 相對的인 크기는 (現生 潮流와 같은 動物들과 比較해 볼 때) 翼龍이 땅 위에서 어떤 生活을 했는지를 보여주는 端緖가 될 수 있다. 아즈다르코類과 翼龍은 몸 크기와 다리 길이에 比해 相對的으로 작은, 아랫다리의 25%-30% 程度 길이밖에 안 되는 발을 가지고 있다. 이것을 보면 아즈다르코類과 翼龍은 相對的으로 단단하고 마른 땅 위를 걷는 데 適應했다는 것을 알 수 있다.
프테라노돈
은 더 큰 발 (
정강뼈
길이의 47%) 을 가지고 있고, 크테노카스마토이드상과(
Ctenochasmatoidea
) 翼龍들과 같이 濾過攝食을 하는 種類는 매우 큰 발 (
프테로닥틸루스
는 정강뼈 길이의 69%, 프테로다우스트로는 84%) 을 가지고 있어서 오늘날의 涉禽類(wading bird)와 類似하게 부드러운 진흙 위를 걷는 데 適應했다는 것을 알 수 있다.
[18]
天敵
[
編輯
]
翼龍은 케찰코아틀루스나 下體고프테릭스같은 大型種을 除外하면 모사사우루스과 爬蟲類나
獸脚類
恐龍의 먹이가 되었던 것으로 알려져 있다. 2004年 7月 1日
네이처
에 古生物學者 에릭 부페타우트는 白堊紀 電氣 地層에서
스피노사우루스
의 부러진 이빨이 박힌 채로 發見된 세 個의 翼龍 경추 化石에 對해 論했다. 이 脊椎뼈들은 하나하나 分離되지 않은 것으로 보아 먹혀서 消化된 것은 아니다.
[41]
生殖과 生活史
[
編輯
]
 졸른호펜 石灰巖에서 發見된 프테로닥틸루스類 새끼의 化石
졸른호펜 石灰巖에서 發見된 프테로닥틸루스類 새끼의 化石
翼龍의 生殖에 對해서는 알려진 것이 거의 없고 알道 가장 稀貴하다. 처음으로 알려진 翼龍 알은 有名한 깃털恐龍들이 發見된 곳과 같은 場所, 랴오닝의 採石場에서 發見되었다. 알들이 깨진 痕跡 없이 납작하게 눌려 있었던 것을 보면 껍질은 가죽 같은 材質로 되어 있었던 것 같다.
[42]
2011年에
다위노프테루스
의 알이 追加로 記載되었는데, 亦是 가죽 같은 材質의 알껍질을 가지고 있어 이 推定을 뒷받침 해주었다. 翼龍의 알은 現生 潮流가 아닌 現生 爬蟲類처럼 어미의 몸 크기에 비하면 매우 작았다.
[43]
翼龍 알 構造와 造成에 對한 硏究가 2007年에 發表되었는데, 이에 依하면 翼龍은 現生
鰐魚
와
거북
처럼 알을 땅 속에 묻었을 갔다고 한다. 알을 땅에 묻는 行爲는 몸무게를 줄이는 適應을 쉽게 만들어 주기 때문에 翼龍의 初期 鎭火에 유리했을 것이지만, 이런 生殖 方法은 翼龍이 살 수 있는 環境의 種類를 一部 制限하며, 後에
鳥類
와 生態學的으로 競爭하게 되는 狀況에서 不利하게 作用했을 수도 있다.
[44]
翼龍 胎兒 化石에서 發見된 飛膜이 잘 發達되어 있는 것을 보면 翼龍은 태어나고 얼마 지나지 않아 날 수 있었던 것으로 보인다.
[45]
태어난 지 며칠에서 一週日 程度 된 翼龍 새끼의 化石도 프테로닥틸루스類, 람포린쿠스류, 크테노카스마투스류, 그리고 아즈다르코類 等 여러 翼龍 科에 걸쳐 發見되었다.
[20]
保存된 뼈들은 모두 새끼들의 나이를 考慮해 보면 相對的으로 骨質化가 相當히 많이 進行되어 있었으며, 날개의 比率은 聖體와 類似했다. 實際로 過去에는 많은 數의 翼龍 새끼들이 聖體로 看做되어 別途의 種으로 分類되곤 했다. 거기에 더해 翼龍 새끼들은 普通 같은 種의 成體 및 未成熟 成長期의 個體과 同一한 場所에서 發見된다. 卽, 프테로닥틸루스와 람포린쿠스의 새끼는 獨逸의 졸른호펜
石灰巖
에서, 프테로다우스트로 새끼는 브라질에서 發見된다. 이들 모두 바닷가에서 멀리 떨어진 깊은 바다 環境에서 發見된다.
[46]
翼龍이 새끼를 돌보았는지는 알려져 있지 않지만 알에서 깨어난 後 얼마 있지 않아 날 수 있었던 것, 그리고 둥지에서 멀리 떨어진 곳에서 聖體와 함께 많은 數의 새끼들이 發見되는 것으로 보아 크리스토퍼 베넷과 데이빗 언윈을 비롯한 大部分의 硏究者들은 새끼들이 父母에 依存하는 時期가 相對的으로 짧았을 것이며, 아마도 날개가 充分히 길어져 혼자 날 수 있게 되기 前, 어쩌면 며칠에 不過했을 빠른 成長期 동안만 보살핌을 받았을 것이며 그 後에는 둥지를 떠나 自立했을 것이라고 結論내리고 있다.
[20]
[47]
다른 可能性으로는, 새끼들이 태어나고 첫 며칠 동안 父母들로부터 먹이를 받아먹는 代身 貯藏되었던 卵黃을 營養分으로 利用했을 수도 있다.
[46]
翼龍의 成長率은 一旦 알에서 깨어나고 나면 種類에 따라 다르다. 람포린쿠스 같은 더 原始的인 긴 꼬리를 가진 翼龍 (람포린쿠스류)의 첫 해 平均 成長率은 130%에서 173% 로,
鰐魚
의 成長率보다 조금 빠르다. 이 종들의 成長은 性的으로 成熟한 以後에는 느려지는데 람포린쿠스가 最大 크기에 到達하기 까지는 3年 以上 걸렸을 것이다.
[47]
이와 對照的으로 더 發展된 大型 프테로닥틸루스類 翼龍들, 例를 들어
프테라노돈
같은 種類는 첫 해에 聖體 크기에 到達한다. 그리고 프테로닥틸루스類는 "恨頂生腸(determinate growth)"을 하기 때문에 定해진 最大 聖體 크기에 到達하면 더 以上 成長하지 않는다.
[46]
一日 行動 패턴
[
編輯
]
翼龍의 一日 行動 패턴을 推論하기 위해 翼龍과 現生 새 및 爬蟲類의 鞏膜고리(sclerotic ring)를 比較하는 方法이 使用되어 왔다. 翼龍 속인
프테로닥틸루스
,
스카포그나투스
, 그리고
투푹수아라
는
晝行性
(diurnal)인 것으로 斟酌되며,
크테爐카스마
,
프테로다우스트로
, 그리고
람포린쿠스
等은
夜行性
,
他페하라
는 하루 全體에 걸쳐 여러 番의 짧은 期間 동안 不規則하게 活動을 했던(cathemeral) 것으로 보인다. 그 結果, 아마도 물고기를 잡아 먹었을 크테爐카스魔와 람포린쿠스는 現生 夜行性 바닷새와 類似한 活動 패턴을 보였을 것으로 보이며 濾過攝食을 했던 프테로다우스트로는 現生 기러기類 中 밤에 攝食을 하는 種類와 비슷한 活動 패턴을 보였을 것이다. 졸른호펜 한 場所에서 發見된 翼龍들인 크테爐카스마, 람포린쿠스, 스카포그나투스, 그리고 프테로닥틸루스의 活動 패턴에 이렇게 差異가 나는 것은 各 속들 사이의
生態地位分割
이었을 수 있다.
[48]
進化와 滅種
[
編輯
]
起源
[
編輯
]
 나무 위의 두 마리
스클레로모클루스
모습 復元도
나무 위의 두 마리
스클레로모클루스
모습 復元도
翼龍의
解剖學
敵 構造가 飛行을 爲해 많이 變形되었고, "잃어버린 고리" 라고 할 수 있는 祖上 形態가 아직 發見되지 않았기 때문에 翼龍의 起源은 거의 잘 理解되어 있지 않다. 몇몇 假說이 있는데,
스클레로모클루스
같은
오르니토디라
와의 聯關性,
에우파케리아
와 같은
支配爬蟲類
와의 聯關性, 或은 프로토社友리아와의 聯關性 等이 提示되었다.
[20]
크리스 베넷(1996)과 데이빗 피터스(2000)는 翼龍이 프로토社友리아와 近緣 關係라고 主張했다. 피터스는 翼龍 化石 寫眞에 寫眞 編輯 소프트웨어의 디지털 트레이싱 機能을 適用하는, DGS 라고 불리는 테크닉을 使用했다.
[49]
베넷은 自身의 分析에서 뒷다리의 特徵들을 除去한 後에야 翼龍과 프로토社우리아가 가까운 關係라는 것을 보일 수 있었다. 이 分析은 翼龍과
恐龍
의 뒷다리에서 볼 수 있는 特徵들이
收斂進化
의 結果라는 생각을 檢證해 보기 위한 試圖였다. 하지만 데이브 魂과 마이클 벤튼의 2007年 硏究에서는 이 結果를 再現할 수 없었다. 魂과 벤튼은 뒷다리의 特徵들 없이도 翼龍과 恐龍이 가까운 關係라는 結果를 얻었다. 이들은 또 데이빗 피터스의 硏究를 批判했는데, 1次 證據, 卽 翼龍 化石을 直接 보지 않고 얻은 結論이 直接 化石을 보고 解釋을 하며 내린 結論과 같은 比重으로 取扱되어야 하는지 疑問을 提起했다.
[50]
魂과 벤튼은 이들의 關係를 分明히 하기 위해 더 原始的인 익룡류의 化石이 必要하긴 하지만 只今까지의 證據들로 볼 때 翼龍은 支配爬蟲類, 特히 오르泥土디라로 看做되는 것이 바람직하다는 結論을 내렸다. 魂과 벤튼의 分析에서 翼龍은 스클레로모클루스의 姊妹그룹이거나 오르泥土디라의 系統圖에서 스클레로모클루스와
라固守쿠스
사이의 位置를 차지하는 것으로 나타났다.
[50]
스털링 네스빗
은 2011年에 스클레로모클루스와 翼龍이 하나의 盆地軍으로 묶일 可能性이 매우 높다는 硏究結果를 發表했다.
[51]
分類
[
編輯
]
系統發生學的 分類學
에서 익룡류(Pterosauria)
盆地群
은 主로 노드에 基盤해 定義되며 몇 個의 잘 알려진
分類群
뿐 아니라 몇몇 原始的인 分類群들에 基盤하고 있다. 2003年의 한 硏究에서는 익룡류를 아누로그나티드과(Anurognathidae),
프레온닥틸루스
, 그리고
퀘찰코아틀루스
의 가장 最近 共通祖上 및 그 모든 後孫을 包含하는 것으로 定義하고 있다.
[52]
이 定義를 따르자면 조금 더 原始的인 近緣種들은 익룡류 盆地軍에 包含되지 않으며 그 代身 더 넓은 盆地軍人 익룡류가
恐龍
보다 翼龍에 가까운 모든
오르니토디라
들을 包含하는 것으로 定義되었다.
[53]
翼龍 內部의
生物學的 分類
는
化石 記錄
에 間隔이 많기 때문에 歷史的으로 어려운 任務였다.이 間隔들이 最近 多量의 새로운 發見에 依해 채워지면서 翼龍의 進化에 對해 더 잘 알게 되었다. 傳統的으로 翼龍은 두 個의
亞目
, 卽 긴 꼬리를 가진 "原始的"인 그룹인
람포린쿠스亞目
과 짧은 꼬리를 지닌 "發展된" 그룹인
프테로닥틸루스亞目
으로 나뉘어 왔다.
[20]
하지만 이 傳統的인 分類는 이제 거의 잘 使用되지 않는다. 프테로닥틸루스類가 람포린쿠스류와의 共通祖上에서 따로 갈라져 나온 것이 아니라 람포린쿠스亞目 안에서 進化한 것이기 때문에 람포린쿠스亞目은
側盆地群
으로 自然스러운 그룹이 아니게 되어
盆地學
李 널리 쓰이게 되면서 大部分의 科學者들이 람포린쿠스亞目이라는 分類를 잘 使用하지 않게 되었다.
[22]
[54]
翼龍들의 正確한 關係는 아직 確定되지 않았다. 翼龍들의 關係에 對한 過去의 많은 硏究들이 一部 制限된 資料만을 包含하고 있었으며 서로 不一致 하는 境遇가 많았다. 하지만 많은 資料들을 使用하는 새로운 硏究들이 이들의 關係를 밝혀가고 있다. 아래의
분지도
는 안드레스와 마이어스의 2013年 硏究에서 提示된
系統發生學
敵 分析을 따른 것이다.
[55]
滅種
[
編輯
]
예전에는 初期
鳥類
種들과의 競爭이 많은 翼龍의
滅種
을 가져왔다고 많이들 생각했다.
[56]
白堊紀 말에 翼龍은 큰 몸집을 가진 種類만 알려져 있다. 작은 종들은 滅種했고, 이들의 生態的 地位는 새들이 차지하게 되었다.
[57]
하지만 翼龍과 鳥類는 生態的으로 겹치는 部分이 別로 없었기 때문에 翼龍의 衰退는 (萬一 衰退가 있었다면 말이지만) 鳥類의 多樣性과는 別 相關이 없었던 것으로 보인다.
[58]
白堊紀가 끝날 무렵,
白堊紀-第3期 大量絶滅
이 非鳥類 恐龍들과 絶對多數의 鳥類 및 많은 動物들을 滅種시켰는데 이 때 翼龍度 滅種한 것으로 보인다. 다른 可能性으로는 一部 翼龍들은 바다에 사는 것에 特化되어 있었던 것으로 보인다. 그 結果 白堊紀-팔레오세 滅種 事件이 翼龍의 먹이가 되는 바다에 사는 生命體들에 影響을 끼쳐 翼龍이 滅種하게 되었을 수 있다. 하지만
아즈다르코과
그리고
이스티오닥틸루스과
는 海洋性이 아니었다.
最近에
픽視
와
오르泥土케이루스
等의 오르泥土케이루스과 翼龍, 프테라노돈과 및 닉土社友루스과, 그리고 타페하라課로 보이는 여러 翼龍들이 캄파니아節/마스트리히트節 連帶의 地層에서 發見되었다.
[55]
[59]
이것을 보면 白堊紀 後期의 翼龍 動物群이 以前에 생각했던 것보다 훨씬 다양했음을 알 수 있으며, 어쩌면 그 多樣性이 白堊紀 電氣에 비해 크게 後退하지 않았던 것일 수도 있다.
프테로닥틸루스類는
白堊紀
電氣의 初期 防産 以後에는 그리 크게 蕃盛하지 않아 그 多樣性이 "墜落"韓 것처럼 보인다. 이 때문에 翼龍의 多樣性은 專門化되어서 작은 滅種 事件에도 脆弱하게 되었을 수 있다.
[60]
그렇지만 아즈다르코과는 白堊紀 末期까지 多樣性이 增加하여 獨特한 生態的 地位를 차지하고 種의 數가 늘었으며 衰退의 徵候는 보이지 않는다.
[61]
잘 알려진 翼龍 속들
[
編輯
]
많이 알려진 翼龍
속
들은 다음과 같다.
- 프테라노돈
은 1.8 미터 程度 몸길이에 날개幅 7.5 미터였고 白堊紀 後期에 살았다.
[62]
- 프테로닥틸루스
는 날개幅 50-75 센티미터였고
쥐라紀
後期에
湖水
가에 살았다.
[63]
- 프테로다우스트로
는 白堊紀의
南아메리카
에 살던 翼龍으로 1.33 미터 程度의 날개幅을 가지고 있었으며 500 個 以上의 길고 가느다란
이빨
을 가지고 있어 이것으로 오늘날의
플라밍고
처럼 濾過攝食을 했을 것이다. 또한, 주된 먹이에 들어있는 色素 때문에 이 翼龍 亦是 플라밍고처럼 粉紅色을 띄었을 可能性이 있다. 南아메리카에서는 처음으로 發見된 翼龍이다.
[64]
- 퀘찰코아틀루스
는 날개幅 10-15미터 程度였고 하늘을 날던 動物 中 가장 큰 축에 屬한다.
[65]
白堊紀 後期에 살았다.
- 오르泥土케이루스
는 白堊紀 電氣인 約 1億 3000萬 年 前에 全世界에 걸쳐 살았던 翼龍이다. 수컷은 부리에 커다란 볏을 가지고 있었다. 過去 最大 翼龍 中 하나였다고 推測되었으나, 硏究 以後 益壯 推定値가 折半假量 줄었다.
- 下體고프테릭스
는 只今까지 알려진 하늘을 나는 動物 中 가장 크다. 全體 骨格 化石이 發見되지는 않았지만
古生物學者
들이 只今까지 찾은 化石을 보면 날개幅은 最小限 10~15 미터에 이를 것으로 보인다.
[66]
- 람포린쿠스
는 쥐라紀에 살던 翼龍으로 꼬리 끝에 날개 같은 것을 가지고 있어서 飛行 中에 꼬리로 움직임을 안정시켰던 것으로 보인다.
[46]
大衆文化 속의 翼龍
[
編輯
]
 王立學會의 350周年을 記念하여 마크 위튼이 製作한
사우스뱅크
의 퀘찰코아틀루스 模型
王立學會의 350周年을 記念하여 마크 위튼이 製作한
사우스뱅크
의 퀘찰코아틀루스 模型
翼龍은 四寸 格인 恐龍만큼 映畫, 文學이나 다른 藝術 形式에서 두드러지지는 않았지만 恐龍만큼이나 오랜 時間 동안 大衆 文化의 한 자리를 차지해 왔다. 게다가 大衆媒體에서 描寫된 恐龍들은 古生物學의 發展에 발맞추어 크게 바뀌어왔지만 20世紀 中盤부터 翼龍의 모습은 繼續해서 時代에 뒤떨어진 狀態로 남아 있었다.
[67]
一般的인 用語인 "프테로닥틸"李 翼龍 全體를 가리키는 單語로 使用되어 왔지만 翼龍의 모습은
프테라노돈
이나
람포린쿠스
, 或은 그 둘이 합쳐진 假想의 모습이 主로 보여졌다.
[67]
아이들 장난감이나 漫畫에서 描寫된 "프테로닥틸"은 "프테라노돈"의 것처럼 생긴 볏과
람포린쿠스
의 것처럼 생긴 긴 꼬리와 이빨로 이루어진 實際로는 全혀 存在한 적이 없던 組合으로 이루어졌다. 最小限 한 種類의 翼龍은 實際로 "프테라노돈"처럼 생긴 볏과 이빨을 가지고 있었다.
屢度닥틸루스
가 그 例인데, 이 이름은 "장난감 손가락"이라는 意味로, 科學的으로 正確하지 않던 아이들 장난감을 닮았기 때문에 붙여진 이름이다.
[68]
또한, 어떤 翼龍 그림은
鳥類
라는 說明이 붙어 있기도 한데, 事實 翼龍은 하늘을 날던
爬蟲類
이며 鳥類는
獸脚類
恐龍의 後孫이다.
翼龍은 아서 코난 도일의 1912年 小說
잃어버린 世界
와 그 後에 發表된 映畫
잃어버린 世界
에서도 登場한다. 以後, 1933年의 映畫
킹콩
, 1966年의 映畫
恐龍 100萬年
等 여러 映畫 및 TV 프로그램에서도 볼 수 있다. 映畫 恐龍 100萬年에서 애니메이터였던
레이 해리하우젠
은 飛膜을 支撐하기 위해 科學的으로는 正確하지 않지만 박쥐의 날개와 같은 形態의 손가락을 스톱모션에 使用된 翼龍 模型에 追加해야 했다. 이 誤謬는 映畫가 만들어지기 前의 그림 等에서도 흔한 것이었다. 翼龍은 2001年의 映畫
쥬라기 公園
3篇 以前까지는 映畫에 그리 자주 登場하지 않았다. 하지만 古生物學者 데이브 魂은 40年이 지났음에도 映畫에 描寫된 翼龍은 現代的인 硏究結果를 反映하지 못하고 있다고 指摘했다. 1960年代부터 2000年代까지 꾸준히 이어진 誤謬들은 이빨이 없는 種類 (쥐라紀 公園 3의 翼龍은 프테라노돈인데 이 이름은 "이빨이 없는 날개" 란 意味이다.) 가 이빨을 가지고 있다거나, 2001年이면 翼龍이 둥지에서 새끼를 돌보지 않는다는 것이 알려졌는데 그런 行動을 보인다거나, 翼龍의 飛行에 必要했고 實際로 翼龍이 가지고 있었던 筋肉纖維로 된 팽팽한 飛膜 代身 펄럭이는 가죽 같은 날개를 가지고 있는 것 等이 있다.
[67]
같이 보기
[
編輯
]
參考 文獻
[
編輯
]
- ↑
Wang X, Kellner AW, Zhou Z, Campos Dde A (2008).
“Discovery of a rare arboreal forest-dwelling flying reptile (
Pterosauria, Pterodactyloidea
) from China”
.
Proc. Natl. Acad. Sci. U.S.A.
105
(6): 1983?7.
doi
:
10.1073/pnas.0707728105
.
PMC
2538868
.
PMID
18268340
.
- ↑
Lawson DA
(March 1975). “Pterosaur from the Latest Cretaceous of West Texas: Discovery of the Largest Flying Creature”.
Science
187
(4180): 947?948.
doi
:
10.1126/science.187.4180.947
.
PMID
17745279
.
- ↑
Buffetaut E, Grigorescu D, Csiki Z (April 2002). “A new giant pterosaur with a robust skull from the latest cretaceous of Romania”.
Naturwissenschaften
89
(4): 180?4.
doi
:
10.1007/s00114-002-0307-1
.
PMID
12061403
.
- ↑
Benton, Michael J.
(2004). “Origin and relationships of Dinosauria”. Weishampel, David B.; Dodson, Peter; and Osmolska, Halszka (eds.).
The Dinosauria
2板. Berkeley: University of California Press. 7?19쪽.
ISBN
0-520-24209-2
.
- ↑
가
나
다
라
Witmer LM, Chatterjee S, Franzosa J, Rowe T (2003). “Neuroanatomy of flying reptiles and implications for flight, posture and behaviour”.
Nature
425
(6961): 950?3.
doi
:
10.1038/nature02048
.
PMID
14586467
.
- ↑
Bennett SC (2000). “Pterosaur flight: the role of actinofibrils in wing function”.
Historical Biology
14
(4): 255?84.
doi
:
10.1080/10292380009380572
.
- ↑
가
나
다
Kellner, A.W.A., Wang, X., Tischlinger, H., Campos, D., Hone, D.W.E. and Meng, X. (2009). "The soft tissue of
Jeholopterus
(Pterosauria, Anurognathidae, Batrachognathinae) and the structure of the pterosaur wing membrane."
Proceedings of the Royal Society B
, published online before print August 5, 2009,
doi
10.1098/rspb.2009.0846
- ↑
가
나
다
라
마
Naish D, Martill DM (2003). “Pterosaurs ? a successful invasion of prehistoric skies”.
Biologist
50
(5): 213?6.
- ↑
가
나
Claessens LP, O'Connor PM, Unwin DM (2009). Sereno, Paul, 編輯.
“Respiratory evolution facilitated the origin of pterosaur flight and aerial gigantism”
.
PLoS ONE
4
(2): e4497.
doi
:
10.1371/journal.pone.0004497
.
PMC
2637988
.
PMID
19223979
.
- ↑
Wilkinson MT, Unwin DM, Ellington CP (2006).
“High lift function of the pteroid bone and forewing of pterosaurs”
.
Proceedings of the Royal Society B
273
(1582): 119?26.
doi
:
10.1098/rspb.2005.3278
.
PMC
1560000
.
PMID
16519243
.
- ↑
Bennett SC (2007). “Articulation and Function of the Pteroid Bone of Pterosaurs”.
Journal of Vertebrate Paleontology
27
(4): 881?91.
doi
:
10.1671/0272-4634(2007)27[881:AAFOTP]2.0.CO;2
.
- ↑
Unwin, D. M.; Frey, E., Martill, D. M., Clarke, J. B., and Riess, J. (1996). “On the nature of the pteroid in pterosaurs”.
Proceedings of the Royal Society B
263
(1366): 45?52.
doi
:
10.1098/rspb.1996.0008
.
- ↑
Wilkinson M.T., Unwin D.M., Ellington C.P. (2006).
“High lift function of the pteroid bone and forewing of pterosaurs”
.
Proceedings of the Royal Society B
273
(1582): 119?126.
doi
:
10.1098/rspb.2005.3278
.
PMC
1560000
.
PMID
16519243
.
- ↑
가
나
Unwin DM, Bakhurina NN (1994). “
Sordes pilosus
and the nature of the pterosaur flight apparatus”.
Nature
371
(6492): 62?4.
doi
:
10.1038/371062a0
.
- ↑
Wang X, Zhou Z, Zhang F, Xu X (2002). “A nearly completely articulated rhamphorhynchoid pterosaur with exceptionally well-preserved wing membranes and "hairs" from Inner Mongolia, northeast China”.
Chinese Science Bulletin
47
(3): 3.
doi
:
10.1360/02tb9054
.
- ↑
Frey, E.; Tischlinger, H.; Buchy, M.-C.; Martill, D. M. (2003). “New specimens of Pterosauria (Reptilia) with soft parts with implications for pterosaurian anatomy and locomotion”.
Geological Society, London, Special Publications
217
: 233.
doi
:
10.1144/GSL.SP.2003.217.01.14
.
- ↑
Dyke GJ, Nudds RL, Rayner JM (July 2006). “Limb disparity and wing shape in pterosaurs”.
J. Evol. Biol.
19
(4): 1339?42.
doi
:
10.1111/j.1420-9101.2006.01096.x
.
PMID
16780534
.
- ↑
가
나
다
라
Witton MP, Naish D (2008). McClain, Craig R., 編輯.
“A reappraisal of azhdarchid pterosaur functional morphology and paleoecology”
.
PLoS ONE
3
(5): e2271.
doi
:
10.1371/journal.pone.0002271
.
PMC
2386974
.
PMID
18509539
.
- ↑
가
나
Frey E, Martill DM (1998). “Soft tissue preservation in a specimen of
Pterodactylus kochi
(Wagner) from the Upper Jurassic of Germany”.
Neues Jahrbuch fur Geologie und Palaontologie, Abhandlungen
210
: 421?41.
- ↑
가
나
다
라
마
바
Unwin, David M. (2006).
The Pterosaurs: From Deep Time
. New York: Pi Press. 246쪽.
ISBN
0-13-146308-X
.
- ↑
Czerkas, S.A., and Ji, Q. (2002). A new rhamphorhynchoid with a headcrest and complex integumentary structures. In: Czerkas, S.J. (Ed.).
Feathered Dinosaurs and the Origin of Flight
. The Dinosaur Museum:Blanding, Utah, 15?41.
ISBN
1-932075-01-1
.
- ↑
가
나
다
라
마
바
Witton, Mark (2013). 《Pterosaurs: Natural History, Evolution, Anatomy》. Princeton University Press. 51쪽.
ISBN
978-0691150611
.
- ↑
Unwin 2005
- ↑
Collini, C A. (1784). "Sur quelques Zoolithes du Cabinet d’Histoire naturelle de S. A. S. E. Palatine & de Baviere, a Mannheim."
Acta Theodoro-Palatinae Mannheim 5 Pars Physica
, pp. 58?103 (1 plate).
- ↑
Wagler, J. (1830).
Naturliches System der Amphibien
Munich, 1830: 1?354.
- ↑
Cuvier G (1801). “[Reptile volant]. In: Extrait d'un ouvrage sur les especes de quadrupedes dont on a trouve les ossemens dans l'interieur de la terre”.
Journal de Physique, de Chimie et d'Histoire Naturelle
52
: 253?267.
- ↑
Arnold, Caroline and Caple, Laurie A. (2004). “Pterodactyl”.
Pterosaurs: rulers of the skies in the dinosaur age
. Houghton Mifflin Harcourt. 23쪽.
ISBN
978-0-618-31354-9
.
- ↑
Naish, Darren.
“Pterosaurs: Myths and Misconceptions”
. Pterosaur.net
. 2011年 6月 18日에 確認함
.
- ↑
Alexander, David E. & Vogel, Steven (2004).
《Nature's Flyers: Birds, Insects, and the Biomechanics of Flight》
. JHU Press. 191쪽.
ISBN
978-0-8018-8059-9
.
- ↑
Alleyne, Richard (2008年 10月 1日).
“Pterodactyls were too heavy to fly, scientist claims”
.
The Telegraph
. 2014年 9月 13日에
原本 文書
에서 保存된 文書
. 2012年 3月 2日에 確認함
.
- ↑
Powell, Devin (2008年 10月 2日).
“Were pterosaurs too big to fly?”
.
NewScientist
. 2012年 3月 2日에 確認함
.
- ↑
Alleyne, Richard (2008年 10月 1日).
“Pterodactyls were too heavy to fly, scientist claims”
. 《The Daily Telegraph》 (London)
. 2015年 8月 20日에 確認함
.
- ↑
Templin, R. J.; Chatterjee, Sankar (2004).
Posture, locomotion, and paleoecology of pterosaurs
. Boulder, Colo: Geological Society of America. 60쪽.
ISBN
0-8137-2376-0
.
- ↑
Naish, Darren
(2009年 2月 18日).
“Pterosaurs breathed in bird-like fashion and had inflatable air sacs in their wings”
. 《
ScienceBlogs
》. 2009年 2月 21日에
原本 文書
에서 保存된 文書
. 2010年 10月 20日에 確認함
.
- ↑
가
나
다
“Why pterosaurs weren't so scary after all”
. 《The Observer newspaper》. 2013年 8月 11日
. 2013年 8月 12日에 確認함
.
- ↑
가
나
Hecht, Jeff (2010年 11月 16日).
“Did giant pterosaurs vault aloft like vampire bats?”
. 《NewScientist》
. 2012年 3月 2日에 確認함
.
- ↑
Padian K (1983). “A Functional Analysis of Flying and Walking in Pterosaurs”.
Paleobiology
9
(3): 218?39.
JSTOR
2400656
.
- ↑
Padian K (2003). “Pterosaur Stance and Gait and the Interpretation of Trackways”. 《Ichnos》
10
(2?4): 115?126.
doi
:
10.1080/10420940390255501
.
- ↑
Hwang K, Huh M, Lockley MG, Unwin DM, Wright JL (2002). “New pterosaur tracks (Pteraichnidae) from the Late Cretaceous Uhangri Formation, southwestern Korea”. 《Geological Magazine》
139
(4): 421?35.
doi
:
10.1017/S0016756802006647
.
- ↑
Unwin DM (1997). “Pterosaur tracks and the terrestrial ability of pterosaurs”.
Lethaia
29
(4): 373?86.
doi
:
10.1111/j.1502-3931.1996.tb01673.x
.
- ↑
Buffetaut E, Martill D, Escuillie F (July 2004). “Pterosaurs as part of a spinosaur diet”.
Nature
430
(6995): 33.
doi
:
10.1038/430033a
.
PMID
15229562
.
- ↑
Ji Q, Ji SA, Cheng YN; 外. (December 2004). “Palaeontology: pterosaur egg with a leathery shell”.
Nature
432
(7017): 572.
doi
:
10.1038/432572a
.
PMID
15577900
.
- ↑
Lu J., Unwin D.M., Deeming D.C., Jin X., Liu Y., Ji Q. (2011). “An egg-adult association, gender, and reproduction in pterosaurs”.
Science
331
(6015): 321?324.
doi
:
10.1126/science.1197323
.
PMID
21252343
.
- ↑
Grellet-Tinner G, Wroe S, Thompson MB, Ji Q (2007). “A note on pterosaur nesting behavior”.
Historical Biology
19
(4): 273?7.
doi
:
10.1080/08912960701189800
.
- ↑
Wang X, Zhou Z (June 2004). “Palaeontology: pterosaur embryo from the Early Cretaceous”.
Nature
429
(6992): 621.
doi
:
10.1038/429621a
.
PMID
15190343
.
- ↑
가
나
다
라
Bennett S. C. (1995). “A statistical study of
Rhamphorhynchus
from the Solnhofen Limestone of Germany: Year-classes of a single large species”.
Journal of Paleontology
69
: 569?580.
JSTOR
1306329
.
- ↑
가
나
Prondvai, E.; Stein, K.; ?si, A.; Sander, M. P. (2012). Soares, Daphne, 編輯.
“Life history of
Rhamphorhynchus
inferred from bone histology and the diversity of pterosaurian growth strategies”
. 《PLoS ONE》
7
(2): e31392.
doi
:
10.1371/journal.pone.0031392
.
PMC
3280310
.
PMID
22355361
.
- ↑
Schmitz, L.; Motani, R. (2011). “Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology”.
Science
332
(6030): 705?8.
doi
:
10.1126/science.1200043
.
PMID
21493820
.
- ↑
Irmis, R. B.; Nesbitt, S. J.; Padian, K.; Smith, N. D.; Turner, A. H.; Woody, D.; Downs, A. (2007). “A Late Triassic Dinosauromorph Assemblage from New Mexico and the Rise of Dinosaurs”.
Science
317
(5836): 358?61.
doi
:
10.1126/science.1143325
.
PMID
17641198
.
- ↑
가
나
Hone D.W.E., Benton M.J. (2007). “An evaluation of the phylogenetic relationships of the pterosaurs to the archosauromorph reptiles”.
Journal of Systematic Palaeontology
5
(4): 465?469.
doi
:
10.1017/S1477201907002064
.
- ↑
Nesbitt, S.J. (2011).
“The early evolution of archosaurs: relationships and the origin of major clades”
(PDF)
.
Bulletin of the American Museum of Natural History
352
: 1?292.
doi
:
10.1206/352.1
. 2019年 7月 1日에
原本 文書
(PDF)
에서 保存된 文書
. 2014年 4月 27日에 確認함
.
- ↑
Kellner, A. W. (2003). "Pterosaur phylogeny and comments on the evolutionary history of the group."
Geological Society, London, Special Publications
,
217
(1): 105-137.
- ↑
Padian, K. (1997). "Pterosauromorpha." Pp. 617-618 in Currie, P.J. and Padian, K.
The Encyclopedia of Dinosaurs
. Academic Press.
- ↑
Lu J., Unwin D.M., Xu L., Zhang X. (2008). “A new azhdarchoid pterosaur from the Lower Cretaceous of China and its implications for pterosaur phylogeny and evolution”.
Naturwissenschaften
95
(9): 891?897.
doi
:
10.1007/s00114-008-0397-5
.
PMID
18509616
.
- ↑
가
나
doi
10.1017/S1755691013000303
現在 이 引用은 內容이 不完全합니다.
英語版의 文書에서 複寫
하여 完成할 수 있습니다.
- ↑
BBC Documentary:
Walking with dinosaurs (episode 4 ) ? Giant Of The Skies
at 22', Tim Haines, 1999
- ↑
Slack KE, Jones CM, Ando T; 外. (June 2006).
“Early penguin fossils, plus mitochondrial genomes, calibrate avian evolution”
.
Mol. Biol. Evol.
23
(6): 1144?55.
doi
:
10.1093/molbev/msj124
.
PMID
16533822
.
- ↑
Butler, Richard J.; Barrett, Paul M.; Nowbath, Stephen and Upchurch, Paul (2009). “Estimating the effects of sampling biases on pterosaur diversity patterns: implications for hypotheses of bird/pterosaur competitive replacement”. 《Paleobiology》
35
(3): 432?446.
doi
:
10.1666/0094-8373-35.3.432
.
- ↑
Agnolin, Federico L. and Varricchio, David (2012).
“Systematic reinterpretation of
Piksi barbarulna
Varricchio, 2002 from the Two Medicine Formation (Upper Cretaceous) of Western USA (Montana) as a pterosaur rather than a bird”
(PDF)
. 《Geodiversitas》
34
(4): 883?894.
doi
:
10.5252/g2012n4a10
. 2013年 1月 15日에
原本 文書
(PDF)
에서 保存된 文書
. 2014年 4月 28日에 確認함
.
- ↑
Wilton, Mark P. (2013). 《Pterosaurs: Natural History, Evolution, Anatomy》. Princeton University Press.
ISBN
0691150613
.
- ↑
Mark Witton, Matyas Vremir, Gareth Dyke, Darren Naish, Stephen Brusatte & Mark Norell. "Pterosaur overlords of Transylvania: short-necked giant azhdarchids in Late Cretaceous Romania" (2013)
- ↑
Wellnhofer, P. (1991).
The Illustrated Encyclopedia of Pterosaurs
. 557?560쪽.
ISBN
0-86101-566-5
.
- ↑
Bennett, S.C. (1996). “Year-classes of pterosaurs from the Solnhofen Limestone of Germany: Taxonomic and Systematic Implications”.
Journal of Vertebrate Paleontology
16
(3): 432?444.
doi
:
10.1080/02724634.1996.10011332
.
- ↑
Palmer, D., 編輯. (1999).
The Marshall Illustrated Encyclopedia of Dinosaurs and Prehistoric Animals
. London: Marshall Editions. 104쪽.
ISBN
1-84028-152-9
.
- ↑
Langston W (1981). “Pterosaurs”.
Scientific American
244
(2): 122?136.
doi
:
10.1038/scientificamerican0281-122
.
- ↑
“Hatzegopteryx”
. 《BBC Nature》. BBC. 2014年 3月 27日에
原本 文書
에서 保存된 文書
. 2012年 3月 2日에 確認함
.
- ↑
가
나
다
Hone, D. (2010).
"Pterosaurs In Popular Culture."
Pterosaur.net
, Accessed 27 August 2010.
- ↑
Frey, E., Martill, D., and Buchy, M. (2003). A new crested ornithocheirid from the Lower Cretaceous of northeastern Brazil and the unusual death of an unusual pterosaur. In: Buffetaut, E., and Mazin, J.-M. (eds.).
Evolution and Palaeobiology of Pterosaurs
.
Geological Society Special Publication
217
: 56?63.
ISBN
1-86239-143-2
.
더 읽기
[
編輯
]
- Witton, Mark (2013).
Pterosaurs: Natural History, Evolution, Anatomy
. Princeton University Press.
ISBN
978-0691150611
.
外部 링크
[
編輯
]