鎭火發生生物學
또는
이보디보
(
英語
:
Evo-devo; Evolutionary Developmental Biology
)는 多樣한
動物
과
植物
의
發生
過程을 比較하여
共通 祖上
에서부터
鎭火
한
生物
의 共通 要素와 變異를 硏究하는
生物學
의 한 分野이다.
鎭火發生生物學
은
生物學
의 한 分野로, 共通의 祖上 關係를 결정짓기 위해 生物들의 發生過程을 比較하고 發生 過程 上의
鎭火
에 對해 硏究한다. 이보디보는
發生
의 根源과 發生 經路의 進化, 發生에서의 變化가 낳은 새로운
形質
들, 發生의 流動性이 進化에서 가지는 役割,
生態學
이 發生的·進化的 變化에 미치는 影響, 進化的 상동의 發生學的 背景에 對해 定義한다. 鎭火發生生物學은 發生의 過程이 어떻게 進化했고,
油田
의 過程에서 어떻게 變更되었으며, 그 結果 어떻게
生物 多樣性
이 形成되는지를 硏究한다.
[1]
槪要
[
編輯
]
發生
過程은
遺傳的
으로 프로그램化되어 있고
鎭火
는 變異와 選擇에 依한 結果이다.
進化生物學
과
發生生物學
은 서로 分離된 關係를 維持하여 오다가 20世紀 初 融合되어
生物 種
사이의 發生 過程 比較가
進化의 證據
로 利用되기 始作하였다. 以後
遺傳學
의 發達로 두 學問 사이의
學際間 硏究
가 繼續되어 왔으며 1980年代 鎭火發生生物學이 成立하게 되었다.
[2]
20世紀 以後 生物學의 硏究 分野는 크게 보아 自然死,
遺傳學
,
發生生物學
으로 區分될 수 있다. 1980年代 後半 始作된 進化發生生物學은 그동안 細分化되어 硏究되었던 生物學의 諸般 硏究 分野를
鎭火
와
發生
을 軸으로 하여 融合한
學際間 硏究
이다. 鎭火發生生物學의 基盤을 이루는 學問은
遺傳學
,
細胞生物學
,
生理學
,
生化學
,
生物物理學
,
內分泌學
等이다.
[3]
槪念 및 意義
[
編輯
]
이보디보란 무엇인가
[
編輯
]
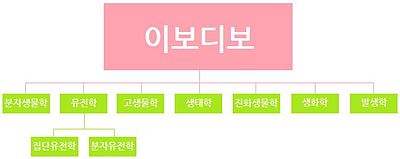 이보디보가 包含하는 學問들
이보디보가 包含하는 學問들
鎭火發生生物學을 가리키는 또 다른 말인 이보디보는 生物學의 한 分野로, 共通祖上 關係를 결정짓기 위해 서로 다른 生物體의 發生過程을 比較하고 發生過程이 어떤 經路로 進化해왔는지에 對해 硏究한다. 이보디보는 發生의 根源과 進化 過程, 發生에서의 變化가 일으킨 새로운
形質
의 誕生, 發生의 流動性이 進化에서 가지는 役割,
生態學
이 發生的?進化的 變化에 미치는 影響, 進化的 상동의 發生學的 背景에 對해 定義한다. 이보디보에서 進化란 發生學的 過程들이 얽혀있는 오래된
遺傳子
體系에서 새롭고 革新的인 構造를 만들거나 다른 生物體들에서 類似한 運動을 하도록 保存하게끔 바꾸는 것이다.
[4]
[5]
學者들의 初期 關心事는 몸의 體制와
組織
의 發生을 調節하는 細胞와 分子水準의 메커니즘에서의 類似點을 發見하는 데에 쏠려있었지만, 現代에는
種 分化
와 聯關된 發生學的 變化들에까지 接近하고 있다.
[6]
이보디보의 意義
[
編輯
]
이보디보는 最近 20年 間 벌어진 生物學의
通涉
흐름을 代辯해주는 學問이라 할 수 있다.
[7]
첫째로, 이보디보는 進化 理論들이 通涉의 길을 걷게 한 張本人이다. 生物學은 1930~1940年代
신다윈주의
나
近代的 綜合
을 한番 겪었으나, 事實 當時의 發生學은 막 登場하기 始作한
遺傳學
에 밀려 生物學界의 統合의 흐름에 함께 끼지 못했다. 遺傳學이
統計
및
確率論
의 도움으로 體系的이고 數學的이라는 評을 받을 때, 發生學은 核心 槪念이었던
形態型成長
(서로 다른 生物들이 멀리 떨어져 있어도 마치 하나의 有機體처럼 情報를 주고 받을 수 있게 만드는 假想의 空間)을 包含하여 難解하고 抽象的인 對象이었다. 따라서 近代的 綜合은 알고 보면 다윈의 自然選擇論과
集團遺傳學
의 만남이었을 뿐이고, 發生學은 그 後 40年이 지나도록 進化論과 統合될 수 없었다. 1970年代 後半에 들어서야
分子生物學
의 엄청난 成功에 힘입어 (分子)
發生遺傳學
者들이 號메오박스를 發見하는 劇的인 成功을 거두면서 어두웠던 發生生物學 硏究에도 햇빛이 들기 始作한다. 이보디보는 以前까지 停滯된 채로 남아있었던 發生學을 눈뜨게 하고, 發生學을
古生物學
및
分子遺傳學
과 結合시키면서 이때까지 풀리지 않았던 問題들을 革命的으로 解決해나간다.
둘째, 이보디보는 進化의 基本原則들을 效率的으로 說明한다. 形態의 進化를 硏究하고, 發生 및 遺傳子에서의 變化가 어떻게 進化를 불러일으키는지 硏究하다 보면, 統一的이면서도 다양한 生物들 아래 숨어있던 原則들이 速速히 드러난다. 또한 이보디보는 進化의 槪念들을 가르치는 데 使用되었던 以前의 抽象的인 接近을 代替하는 具體的이고 視覺的인 接近法들을 提案한다. 例를 들어, 胚芽에서 遺傳子가 發現하는 形態는 視覺的으로 뚜렷하며, 서로 다른 種의
툴킷 遺傳子
들은 各各을 집어내 보일 수 있는 것으로 훨씬 效率的으로 進化의 槪念들을 說明할 수 있다.
셋째, 이보디보는
進化生物學
敎育을 두고 빚어지는 社會的 論難에서 큰 도움이 될 수 있다. 이보디보는 進化의 메커니즘과 原則들을 實體的이고 具體的으로 說明하기 때문이다.
마지막으로, 鎭火發生生物學의 重要性은 確實히
哲學
的인 水準을 넘어선다. 우리 周邊의 生態系는 人間뿐만 아니라 無數히 다양한 生命體들로 이루어져 있다. 自然 안에서 우리는 모두 相互作用하며 서로에게 至大한 影響을 끼친다. 따라서, 모든 生命體들의 運命은 우리 人間이 進化에 어떤 影響을 미치는가를 먼저 理解할 때 保障받을 수 있다.
이보디보의 歷史 및 背景
[
編輯
]
이보디보의 歷史
[
編輯
]
1850年
오웬
은 生命의 歷史가 目的을 가진 段階的인 計劃에 따라서 自然的인 發生에 依하여 만들어진
種
들에 依하여 새로운 種들이 만들어지며 運命的으로 쓰인 것이라고 主張하였다.
[8]
[9]
찰스 다윈
銀 種의 起源에서
自然選擇
을 통한
鎭火
를 主張하여 현대
生物學
의 기둥을 세웠다. 그는 일찍이 進化를 硏究하는 데 있어서
發生
의 重要性을 認定하였다.
[10]
| “
|
내가 보기에 發生學은 形態의 變化를 支持하는 가장 强力한 事實들이 담긴 學問인데, 내 冊을 評하는 사람들 中 그 點을 言及하는 사람은 내가 보기에 아무도 없습니다. ? 찰스 다윈, 餓死 그레이에게 보낸 便紙에서, 1860年 9月 10日
|
”
|
또한 다윈은
폰 베어
의
盆地形態理論
과 그가 主張한 變化를 隨伴한 後孫의 槪念을 連結 지어서 왜 自然的 分類를 할 때,
胚芽
로부터 誘導된 特徵들이
聖體
로부터 誘導된 特徵들과 同等한 重要性을 가지는지에 對한 解答을 내놓았다.
에른스트 헤켈
은
라마르크 注意
와
自然哲學
을 統合하여 다윈의 綜合說을 만들어내려고 努力했다. 그는 ‘個體發生이 系統發生을 再演한다’라는 主張을 하였다.
[11]
卽, 모든 種의 胚芽의 發生(個體發生)만으로 그 種의 進化的 發生過程(系統發生)을 說明할 수 있다는 것이다.
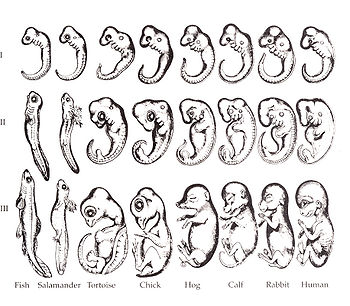 1874年 헤켈은 여덟 種의 脊椎動物―魚類, 도롱뇽, 거북이, 닭, 돼지, 소, 토끼, 사람―의 胚芽의 發生 過程을 그렸다.
1874年 헤켈은 여덟 種의 脊椎動物―魚類, 도롱뇽, 거북이, 닭, 돼지, 소, 토끼, 사람―의 胚芽의 發生 過程을 그렸다.
當時 그의 理論은 大部分의 사람들의 信任을 얻지는 못했지만, 현대 進化論 綜合설이 세워진 以來 發生에 對한 새로운 關心을 불러 일으키는 契機가 되었다.
스티븐 제이 굴드
는 이 接近을
宗團附加
로 進化를 說明하는 데 利用하였다. 宗團附加란 個體 發生의 마지막 過程에 새로운 形質이 더해지는 것을 말하는데, 이는 個體發生이 系統發生을 再演한다는 헤켈의 主張을 뒷받침하기 위하여 헤켈主義者(或은
反復론자
라고 한다)들이 만들어낸 槪念이다. 헤켈의 主張이 說明되기 위해서는 系統 發生의 過程에서 새로운 形質이 恒常 祖上 生物의 個體 發生 途中이 아닌 마지막에 더해져야 하기 때문이었다. 그러나 이러한 主張은 宗團附加가 일어나면 祖上 生物에 비해 後孫 生物의 個體 發生이 차츰 길어질 수 밖에 없다는 問題를 가지고 있었다. 이를 說明하기 위하여 反復論者들은 새로운 段階가 더해지며 進化的 發展이 일어나면, 恒常 오래된 段階를 壓縮하거나 削除하는 過程이 일어나 發生 時間이 길어지는 것을 막는다고 說明했다. 이 主張은 어느 瞬間 動物의 成長이 遲延되고 生殖器官이 成熟하여 繁殖하는 形象, 卽
幼形成熟
을 觀察한 것에 基盤을 두고 있었다.
[12]
幼形成熟의 發見은 個體 發生에서 어떤 器官의 發生이 特히 促進되거나 遲延되는
이시성
의 觀點에서 擴張되어, 進化的 變化의 메커니즘으로 받아들여지게 되었다.
[13]
다아시 톰슨
은 1917年에 出版한 그의 冊 『成長과 形成』에서 成長 速度의 差異가 形態 上의 變異들을 만든다고 宣言했다. 그는 生物 個體의 體制에 있어서 根本的인 類似點들과 幾何學的인 變異들이 생기는 過程이 變異들을 說明하는 데 있어서 利用될 수 있다고 보였다.
1970年代 後半
獨逸
의 生物學者인
에드워드 루이스
等은
노랑초파리
에서 180個의
鹽基序列
로 構成된 特定
DNA
短篇인
號메오박스
를 發見하였다. 1980年代에 들어 同一한 鹽基序列을 갖는
號메오박스
가
哺乳類
에서도 發見되었으며 이들이 同一한 役割을 한다는 것이 밝혀졌다. 醋파리 胚芽의 앞과 뒤를 決定하는 號메오박스 遺傳子는 哺乳類에서
脊椎
形成에 關與한다. 卽, 類似한 遺傳子는
系統
敵으로 멀리 떨어져 있는 生物에서도 類似한 役割을 한다는 點이 밝혀진 것이다. 醋파리의 特定 號메오박스 遺傳子를 생쥐의
胚芽
에 移植한 實驗에서 醋파리의 號메오박스 遺傳子는 正常的인 생쥐의 그것과 同一한 役割을 遂行하였다. 號메오박스의 發見은 胚의 發生과 進化에 對한 새로운 關心을 불러일으켰고 鎭火發生生物學이 始作되는 契機가 되었다. 號메오박스의 發見은 胚의 發生과 進化에 對한 새로운 關心을 불러일으켰고 鎭火發生生物學이 始作되는 契機가 되었다. 1995年 루이스와 同僚들은 號메오박스 發見의 功勞로
노벨 生理學·醫學賞
을 受賞하였다.
[3]
1988年
리보솜 RNA
의 鹽基 序列을 比較하여
動物界
全體의 祖上 後孫 關係를 把握하는 硏究가 進行된 以後 DNA의 鹽基 序列을 比較하여 生物의
系統
을 밝히는 分子系通學 硏究가 始作되었다. 現在 모든 動物 系統의 盆地가 밝혀지지는 않았으나 分子系統學의 硏究는 比較生物學의
相同性
硏究에 많은 도움을 주고 있다.
[2]
硏究
[
編輯
]
進化生物學의 硏究는
사람
과 醋파리 사이에 共通的인 遺傳子는 어떤 것이 있는 지, 또한 사람만이 갖는 固有한 遺傳子에는 어떤 것이 있는지를 硏究한다. 比喩하자면 號메오박스와 같은 遺傳子는 서로 다른 模樣을 하고 있는
레고
블록과 같다. 같은 種類의 레고 블록을 使用해서 집을 만들 수도 있고 自動車를 만들 수도 있듯이, 서로 다른 特徵을 갖는 遺傳子들이 모여서 한 生物의
遺傳體
를 이루게 된다. 進化의 긴 歷史 속에서 어떤 遺傳子는 生物 全體에 걸쳐 共通的으로 남아 있을 수도 있고 다른 것은 빠지거나 바뀔 수도 있다. 이렇게 生物種의 發生 特徵을 결정짓는 遺傳子를 調節部位라고 한다. 사람과 醋파리에게는 앞서 보았던 共通的인 役割을 하는 調節部位度 있는 反面,
턱
과 같은 骨格을 形成하는 調節部位는 醋파리에게는 없다.
[1]
號메오 遺傳子(혹스 遺傳子, 體質決定遺傳子
[14]
)
[
編輯
]
號메오박스
는 藥 180個의
鹽基
雙(
뉴클레오타이드
)으로 이루어져 있으며, 수많은 遺傳子들의 스위치를 켜는
戰士人者
들을 暗號化한다. 號메오박스 遺傳子는 發現되어 序列 特異的으로 DNA와 結合할 수 있는
號메오도메인
이라고 불리는 蛋白質을 만든다. 醋파리의 이런 遺傳子 가운데 하나에 突然變異가 생기면 더듬이 位置에 다리가 생긴다. 아래의 序列은 典型的인
인트론
들을 대시記號로 標示한 號메오박스 領域에 該當하는 60個의
폴리펩타이드
사슬을 보여준다.
[15]
RRRKRTA-YTRYQLLE-LEKEFLF-NRYLTRRRRIELAHSL-NLTERHIKIWFQN-RRMK-WKKEN
 HoxD 遺傳子무리
HoxD 遺傳子무리
하나의 號메오도메인 蛋白質만으로는 타깃이 되는 遺傳子만을 認識해내기가 힘들기 때문에, 大部分의 境遇 號메오도메인 蛋白質들은 다른 戰士因子들과 함께 複合體를 이루며 그들의 타깃 遺傳子들의
프로모터
部位에서 役割을 遂行한다. 이러한 複合體들은 單一 號메오도메인 蛋白質보다 훨씬 높은 타깃 特異性을 보인다.
最初로 號메오도메인을 찾은 곳은 醋파리의
號메오蛋白質
들과
分節形成蛋白質
들이었다. 以後
發生遺傳學
者들의 刻苦의 努力으로 脊椎動物을 包含한 많은 動物들에서 號메오도메인이 保存되어 있음을 밝혔다.
[16]
[17]
[18]
 안테나페디아 突然變異가 생긴 醋파리의 머리
파일:Bithorax.jpg
바이土락스 突然變異가 생긴 醋파리
안테나페디아 突然變異가 생긴 醋파리의 머리
파일:Bithorax.jpg
바이土락스 突然變異가 생긴 醋파리
號메오도메인은
나선 臺 羅先連結構造
를 만들며 DNA와 結合한다. 나선 臺 나선연결구조는 두 個의
알파螺線
들로 이루어진 特徵을 지니는데, 이 나선들은 DNA와 號메오도메인이 가까이 닿게 하고 짧은 回戰 동안에 結合할 수 있게 한다. 이中 두 番째 螺旋은
곁사슬
들과 DNA의 回轉으로 생기는
駐홈
에서 밖으로 露出된 鹽基들과
티민
메틸기
들 사이의 水素結合과
疏水性
의 相互作用들을 통해 DNA와 結合한다.
[18]
DNA와 號메오도메인의 두 番째 螺旋이 結合하면, 첫 番째 螺旋은 그 構造의 安定化를 돕는다.
號메오 遺傳子들은 胚芽의
前後軸
을 따라서 胚芽 內 各 區域의 運命을 결정짓는 데 必須的인 遺傳子다. 號메오 遺傳子들은 號메오도메인을 包含한
戰士調節因子
들을 暗號化하는데, 戰士調節因子들은 動物의 前後 體軸을 따라서 다른 遺傳的 프로그램을 作動시킨다.
[19]
1984年에
에디 드 로버티스
는
손톱개구리
속에서 最初로
脊椎動物
의
號메오 遺傳子
를 分離하여 얻어, 이보디보 發展의 原動力을 提供했다.
[20]
脊椎動物에서, 같은 祖上 遺傳子에서 誘導된 네 個의 遺傳子들은 若干의 機能을 喪失했지만, 同時에 여러 가지의 새로운 機能들을 獲得했다. 例를 들어,
HoxA
와
HoxD
遺傳子는 다리 軸을 따라 그 區域의 運命을 결정짓는다.
이러한 遺傳子들의 주된 魅力은 바로 그들의 獨特한 行動에 있다. 號메오 遺傳子들은 普通 組織化된 하나의 무리에서 發見된다. 무리 內에서 이 遺傳子들의 配列 順序는 影響을 끼치게 되는 區域과 時期와 直接的으로 聯關되며, 이러한 現象을
共通直線性
理라 한다. 共通直線性으로 因해, 突然變異로 인한 遺傳子 무리의 變化는 普通 遺傳子의 影響이 미치는 區域에 類似한 變化를 일으킨다.
例를 들어, 動物의 特定 部分이 動物의
全體富
로 發生하는 것을 擔當하는 한 遺傳子에
突然變異
가 생기면 그 部分이 元來보다
後體部
에 發生하게 된다. 이 現象을
前衛
라 한다. 醋파리에서의 前衛의 有名한 例示로는, 머리에 더듬이 代身 다리가 생기는
안테나페디아
와 두 個의 가슴이 생기는
바이오토락스
突然變異가 있다.
分子的 證據들은 몇 個의
號메오 遺傳子
들이
左右對稱動物
以前의
刺胞動物
에서도 發見됨을 보여주는데, 이것으로 號메오 遺傳子들이
古生代
以前부터 存在했다고 推定된다.
[21]
遺傳子 툴킷
[
編輯
]
마스터 遺傳子
中 가장 有名한 것들인 號메오 遺傳子, 눈, 附屬肢, 心臟을 만드는 十餘 個의 遺傳子들도 動物 發生을 책임지는 遺傳子 툴킷 가운데 一部分에 不過하다. 醋파리 形成에 關聯된 遺傳子만 해도 數百 個 程度이고, 나머지 수많은 遺傳子들도 各自 醋파리 細胞들 안에서 特化된 機能을 遂行하는 일을 한다. 또한 動物 身體의 發生과 무늬 形成을 統制하는 動物 發生의 툴킷에는 號메오 蛋白質, 身體 形成 마스터 遺傳子들, 細胞
形態調節因子
들,
헤지호그
및 其他
信號傳達 蛋白質
들, 細胞
受容體
들,
호르몬
들,
着色 蛋白質
들, 기타
DNA 結合 蛋白質
들이 있다.
[22]
遺傳學者들은 全體 툴킷 遺傳子를 밝혀내기 위해서 畸形을 띠는 突然變異體의 遺傳子들을 分離한 後 分析하였다. 1970年代 末부터 1980年代 初에
크리스티안네 뉘슬라인-폴하르트
와
에릭 位샤우스
는 醋파리
幼蟲
이 發達하는데 必要한 거의 모든 遺傳子들, 卽 適切한
體節
을 만드는 데 必要한 數十 個의 遺傳子들, 세 겹의
組織層
을 만드는데 必要한 遺傳子들, 그 밖의 裝飾에 必要한 遺傳子들을 낱낱이 밝혀냈다.
[23]
甚至於 醋파리에 對한 硏究를 擴張시켜 大部分의 脊椎動物과 다른 動物도 이와 類似한 遺傳子들을 가지고 있다는 것을 알아내면서, 이보디보의 發展의 土臺를 닦았다.
[24]
그런데 뉘슬라인-폴하르트와 位샤우스가 發見해 온 突然變異들에게서 보인 衝擊的인 共通點이 있었다. 바로, 遺傳子들이 昆蟲 構造의 基礎
모듈
이 되는 體節 單位 以上으로만 影響을 미쳤다는 點이다.
[25]
[26]
遺傳子들은 基礎가 되는 體節의 救助는 保存하지만, 體節의 配列에만 影響을 미쳤다. 德分에 어떤 境遇라도 發生이 마구잡이로 되지는 않았고, 特定 體節들이 全部 或은 一部가 없어지거나 極盛이 흐트러지는 패턴의 變化를 나타냈다. 또한 特定 機能이 비활성화되어도 다른 機能들은 正常的으로 作動했다.
以後 툴킷 遺傳子들에 對한 많은 硏究 끝에, 오늘날 툴킷의 수많은 遺傳子들이 알려져 있다. 툴킷 遺傳子들은 大體로 다른 遺傳子들에 對한 스위치 役割을 해서 發生에 影響을 미친다. 툴킷의 큰 部分을 차지하는 것으로는
戰死 要因
과
信號傳達經路
에 屬하는 蛋白質이 있다. 戰死 要因은,
마스터 遺傳子
와 같이 DNA에 直接 結合해서 遺傳子의
戰死
를 껐다 켰다 한다. 한便, 한 細胞가 다른 細胞로 疏通하기 爲해서 分泌한
信號 蛋白質
은 다른 細胞의
受容體
에 結合해서 그 細胞의 模樣과
分列
等에 關한 여러 變化를 일으킨다. 醋파리 內에도 約 열 가지의 信號傳達經路가 있는데, 細胞와 細胞를 넘나드는 信號, 受容體, 갖가지
中間物質
等의 信號傳達 要素들 中 하나라도 突然變異가 일어나면 全體 信號 體系가 무너지게 된다. 以後 生物學者들은 脊椎動物에서 醋파리의 툴킷 遺傳子와 上洞人 遺傳子를 찾아냈다.
[27]
윌리스턴의 法則
[
編輯
]
古生物學者
새뮤얼 윌리스턴
은 1914年 古代
海洋 爬蟲類
를 硏究하던 途中 初期 動物群에는 비슷한 附屬들이 多數 反復되는 反面, 後代의 動物群에서는 附屬의 數가 줄고 救助마다 한결 專門化된 形態가 나타난다는 事實을 알아차렸다. 그는 이 事實에서 ‘鎭火는
有機體
身體附屬들의 數가 줄어드는 方向으로, 줄어든 部位들이 機能 面에서는 훨씬 專門化되는 方向으로 進化한다’라는 윌리스턴의 法則을 導出하였다. 쉽게 말해서 充分한 數를 確保한 連續
相同器官
들은 機能의 專門化와 數의 減少를 向해 간다는 것이다.
[28]
醋파리에서의 號메오 遺傳子
[
編輯
]
지난 世紀부터
슈페만
,
베이트슨
과 같은 生物學者들은 自然과 實驗 途中에 생긴 怪物들의 誕生에 對해 疑問을 가졌다. 그들은 이러한 怪物들을 通해서
大腸菌
에서부터 코끼리까지 모든 生物에서 통할 수 있는 發生의 規則을 알아내기 위해 끊임없이 努力했다.
[29]
[30]
이 發生의 法則을 밝히기 위해서는 正常과 다른 畸形들을 數없이 確保하고, 突然變異 形質을 純種으로 길러 後孫들이 同一한 特性을 나타냄을 보여야 했다. 1915年, 유전학자
캘빈 브리지스
가 偶然히 작은 뒷날개가 앞날개의 크기만큼 큰 醋파리 突然變異體를 發見한 以後로
[31]
, 다리가 머리에 달린
안테나페디아
突然變異 等의 여러
號메오 突然變異
가 醋파리에서 發見되면서 醋파리에서의 이보디보 硏究가 本格的으로 물꼬를 틀기 始作했다.
[32]
[33]
醋파리에서의 號메오 突然變異體들의 魅力은 單 하나의 遺傳子의 問題로 몸 全體가 全혀 다르게 變했다는 것에 있었다. 이 問題의 解答을 찾는 데 열쇠가 되는
遺傳子 複製
技術이 普遍化되면서, 몇몇 生物學者들이 醋파리 突然變異에 對한 硏究에 뛰어들었다. 그들은 數年의 硏究를 통해서 醋파리의 號메오 遺傳子들이 두 個의 複合體로 나뉘어서, 醋파리의 總 네 染色體 中 세 番째 染色體에 位置한다는 것을 알았다. 첫 番째 複合體는 세 個의 遺傳子로 이루어져 있고 醋파리의
全體富
를, 두 番째 複合體는 다섯 個의 遺傳子로 이루어져 있고 醋파리의
後體部
를 調節하고 있었다. 신기한 點은 바로 號메오 遺傳子들이 複合體 속에 配置된 順序가 遺傳子가 調節하는 몸 部位의 順序와 一致한다는 것이었다.
[34]
1983年 以來 生物學者들은 두 複合體를 分析하면 醋파리 形成의 規則을 찾아낼 수 있다는 期待 아래에서 두 複合體의 DNA를 分析하기 始作했다. 硏究 結果, 複合體 內 8個의 遺傳子로부터 다양한 號메오 蛋白質들이 發現되고, 이들은 各各 特定 身體部位에 獨自的인 影響을 미치면서 한便으로는 約 아미노산 60個에 該當하는 類似序列을 지니고 있어 共通的인 機能을 遂行하고 있었다. 號메오 遺傳子들에 共通的으로 存在하는 이 180個 鹽基雙 序列은 DNA 內에서 箱子 模樣으로 보였으므로 號메오박스 遺傳子라 불렸다.
[35]
[36]
이 같은 놀라운 發見에도 不拘하고, 當時 生物學界의 定說에서는
哺乳類
와 醋파리와 같은 벌레는 發生法則 自體가 매우 다르다고 생각했으므로 크게 인정받지 못했다. 哺乳類를 硏究하는 生物學者와 比較的
何等生物
들을 硏究하는 生物學者들의 交流가 없어 일어난 일이었다. 例外的으로
빌 脈기니스
와
마이크 레빈
은 硏究 途中 號메오 突然變異에 마음을 뺏기고, 哺乳類가 특별하다는 以前의 偏見을 버렸다. 두 사람은 醋파리의 모든 號메오 遺傳子들에 號메오박스가 있다는 것을 確認하고, 周邊에서 찾을 수 있는 벌레, 지렁이, 개구리, 蘇, 사람 等과 같은 온갖 生物들에서 DNA를 分離해 號메오박스를 찾기 始作했다. 놀랍게도 두 사람은 醋파리 以外의 動物에서도 號메오박스들을 發見하고 甚至於 그것들이 매우 類似한 序列을 가지고 있음을 알아냈다.
[37]
이를 通해, 學者들은 號메오 遺傳子들은 모든 生物의 發生에서 核心的인 役割을 하므로, 잘 保全되어 왔음을 推定할 수 있었다.
몇 年이 지난 後, 쥐의 號메오 遺傳子 配列을 醋파리와 比較해본 結果 마찬가지로 遺傳子들이 몇 個로 나뉜 複合體로 存在하였다. 뿐만 아니라, 各 複合體 속 遺傳子 配列은 醋파리에서와 같이 遺傳子가 發現되어 影響을 미치는 쥐의 身體部位 順序와 一致했다.
[38]
[39]
이 硏究를 통해, 號메오 遺傳子 複合體들이 파리와 쥐처럼 서로 다른 動物들의 發生에서 遺傳子의 鹽基 序列, 遺傳子들이 複合體로 存在하는 方式, 甚至於는 胚芽에서 利用되는 方式에 같은 影響을 미치고 있음을 否定할 수 없게 되면서, 生物界에 巨大한 革命의 바람이 몰려올 것을 暗示하게 되었다.
또한 빌 脈기니스와 마이크 레빈이 있었던
발터 게링
實驗室에서는 醋파리의
아이리스
(눈 없음) 遺傳子를 發見하고 사람에게도 이와 對應되는 遺傳子
아니리디아(무홍채)
가 있다는 것을 알았다.
[40]
아니리디아 遺傳子는 또한 쥐의
스몰아이
遺傳子와 같았다. 相異한 눈 組織을 形成하는 데 같은 遺傳子가 關聯됨에 對해 疑問을 가진 硏究者들은 追加로 한 實驗에서, 아이리스 遺傳子가 醋파리의 다른 身體部位에서 發現되도록 造作을 하면 그 部位들에서 눈 組織이 形成될 수 있음을 確認했다.
[41]
이로써 아이리스 遺傳子가 눈 發生을 調節하는
마스터 遺傳子
임을 알게 되었다. 더 놀라운 結果는 쥐의 스몰아이 遺傳子를 醋파리에 넣었을 때, 쥐가 아닌 醋파리의 눈 組織이 誘導되었다는 것이다.
쥐 遺傳子가 파리의 눈 發生을 促進한 것이었다. 以後, 아이리스, 아니리디아, 스몰아이 遺傳子를 묶어
팍스-6
遺傳子라 부르기 始作했고, 이들은 모든 種類의 動物에서 눈 形成과 聯關되어 있음으로 보아 모든 生物의 共通 祖上이 팍스-6 遺傳子를 눈 形成에 使用했으므로 進化되는 내내 再使用되고 保存되어왔음을 推定할 수 있었다. 또 다른 例로, 遺傳子에 損傷을 입으면 醋파리의 다리의 바깥쪽에 變異가 일어나는
디스탈리스
遺傳子가 모든 生物의
附屬肢
를 만드는데 使用되고
[42]
, 醋파리의 心臟 形成에 必須的인
틴먼
遺傳子들이 脊椎動物의 心臟 形成에도 重要하게 作用한다고 밝혀졌다.
[43]
些少한 醋파리의 號메오 突然變異를 始作으로, 오랜 期間 維持되어왔던 進化生物學者들의 ‘서로 다른 動物들은 오랜 期間 동안 各自 다른 進化의 歷史를 거치면서 完全히 別個의 方法으로 進化되어 왔다’는 생각은 무너졌다. 相異한 動物에서 共通된 遺傳子가 動物의 發生에 매우 類似한 機能을 함이 알려지자, 生物學者들은 다시 動物의 發生과 構造의 起源, 動物의 進化에 對해 徹底히 再檢討해야 하는 立場에 處하게 된다.
이보디보의 擴張
[
編輯
]
 遺傳子에 突然變異가 생겼을 境遇 사람에서 多指症을 일으키는 것으로 알려진 헤지호그 遺傳子가 병아리에서도 同一하게 作用하며, 醋파리에서는 醋파리 幼蟲에 많은 纖毛가 돋아나게 한다.
遺傳子에 突然變異가 생겼을 境遇 사람에서 多指症을 일으키는 것으로 알려진 헤지호그 遺傳子가 병아리에서도 同一하게 作用하며, 醋파리에서는 醋파리 幼蟲에 많은 纖毛가 돋아나게 한다.
醋파리에서 號메오 遺傳子를 發見한 後,
分子生物學
者들은 醋파리의 號메오 遺傳子를
探針
으로 使用하여 다른 生物에도 이러한 遺傳子가 存在하는지를 알아보는 作業에 着手했다. 結果는 大成功으로,
船蟲
에서 코끼리까지 모든 動物에서 號메오 遺傳子의 存在가 確認되었다. 甚至於 鹽基 序列 또한 매우 비슷함을 보였다.
[37]
이는 號메오박스가
眞核生物
의 進化 過程에서 매우 初期에 일어났음을 示唆한다. 쥐의 號메오 遺傳子의 配列에 關한 硏究는 號메오박스의 機能에 疑懼心을 품고 있던 學者들 조차 그 重要性을 認定하지 않을 수 없게 하였다. 쥐의 號메오 遺傳子들은 파리와 마찬가지로 몇 個의 複合體를 이루고 있었으며, 甚至於 그 順序는 發現되는 쥐의 身體部位 順序에 正確하게 對應했다. 이는 動物들의 號메오 遺傳子의 類似性이 序列의 類似性으로 그치는 것이 아니라 複合體 組織을 이루는 方式, 나아가 胚芽에서 活用되는 方式에까지 미친다는 것을 보여준다. 이로써 號메오 遺傳子 複合體들이 動物들의 發生에 同一하게 影響을 미치고 있다는 事實은 否認할 수 없게 되어 버렸다.
[38]
身體 形成 遺傳子 툴킷
[
編輯
]
어떻게 醋파리와 사람처럼 確然하게 다른 身體 構造를 形成하는 遺傳子의 序列이 그토록 비슷할 수 있는 것일까? 처음에 科學者들은 醋파리와 사람의 너무나도 큰 解剖學的 構造 差異 때문에 醋파리의 發生學的 過程 硏究가 사람의 發生學的 過程의 硏究에 아무런 도움도 주지 못할 것이라고 생각했다. 그러나 곧 이것은 틀린 생각이라는 것이 證明되었다. 醋파리의 눈을 形成하는데 關聯하는 마스터 遺傳子인 '아이리스 遺傳子'와 對應되는 遺傳子 軍이 다른
생물군
에서도 發見되었고, 甚至於 다른 生物群의 눈-形成
마스터 遺傳子
를 醋파리에 집어넣어도 正常的인 醋파리의 눈이 形成되었다.
[41]
또한 醋파리의
末端部
를 形成하는 遺傳子인
디스탈리스
遺傳子 또한 醋파리와 類緣關係가 적은 動物을 包含한 모든 動物의 몸에서 튀어나온 附屬肢들의 形成에 모두 關聯됨을 보였다.
[42]
共通의 툴킷 遺傳子
[
編輯
]
크리스티안네 뉘슬라인-폴하르트
와
에릭 位샤우스
의 硏究에 依하여 醋파리 幼蟲의 發達에 必要한 모든 遺傳子들이 알려지고
[23]
, 大多數의 脊椎動物과 其他 動物들이 이에 對應하는 遺傳子들을 갖고 있다는 事實이 밝혀진 以後, 툴킷 遺傳子의 突然變異는
모듈
式 構造는 維持한 채 配列의 패턴에만 影響을 미친다는 事實이 밝혀졌다. 툴킷 遺傳子들은 다른 遺傳子들의 戰史에 關한 스위치를 調節하며 發生에 影響을 주는데,
戰士人者
는 이러한 툴킷 遺傳子의 큰 部分을 차지한다.
[44]
툴킷 逆說과 多樣性의 起源
[
編輯
]
共通된 툴킷은 툴킷 遺傳子가 매우 오래된 것으로서 大部分의 動物들이 갈라져 進化하기 前부터 存在했다는 事實을 의미한다. 사람과 醋파리처럼 서로 相異하게 다른 形態를 가진 動物이라도 共通의 툴킷을 가지고 있을 뿐만 아니라
遺傳體
의 序列 또한 매우 恰似하다. 이러한 事實로부터 共有하는 遺傳子가 이렇게나 많은데 어떻게 그렇게 相異한 形態가 만들어지는지에 對한 逆說을 提起할 수 있다. 그 解答은 바로 툴킷의 스위치 調節에 있다.
[45]
動物 多樣性 形成
[
編輯
]
一連의
發生學
敵 硏究를 통해 科學者들은
現生 生物
의 툴킷 遺傳子를 밝혀 내고 그 役割과 모든 動物에서 툴킷의 共通性 또한 밝혀낼 수 있었다. 이보디보는 이에서 그치지 않고 過去의 生物에서 어떻게 只今의 다양한 形態의 生物群이 形成되었는지를 밝혀냄으로써 進化의 現代的 綜合 理論의 큰 寄與를 하였다. 이보디보가 主張하는 進化的 革新의 主要 原理는 이미 存在하는 構造가 變形되어 새롭게 進化된 構造가 形成된다는 것이다. 進化的 革新으로 가는 가장 쉬운 길은 一旦 A로 간 後에 B로 가는 것이지, 아무것도 없는 곳에서 곧바로 B로 가는 길을 말하는 것이 아니라고 이보디보는 말한다. 또 하나의 原理는
多技能性
과 重複性이다. 여러 機能을 擔當하는 構造가 있는데 그 構造가 여러 個 重複되어 存在한다면, 勞動 分業을 이루어 서로 다른 構造로 專門化되어 다양한 機關을 만들 수 있다. 마지막으로
모듈性
이 있다.
翼龍
이 네 番째 손가락을 길게 進化시키고, 뱀이 數百 個의
脊椎
뼈들을 進化시켜 몸통을 늘이고,
박쥐
의 손가락들이 길어져 날개막을 支持할 수 있었던 것은 모두 이들이 모듈 式 設計를 取하고 있었기 때문이다. 모듈性 德分에 各 身體 附屬들은 다른 附屬들에게 全혀 影響을 미치지 않고 獨立的으로 變形되거나 專門化 될 수 있었다. 이보디보의 論理에 따르면 生物이 이토록 다양한 形態를 띠게 된 것은 自明하다. 動物은 革新을 이루어 새로운
生態地位
를 얻을 수 있었고, 새로운 生態地位는 또한
多樣性
의 擴張을 促進했다
[46]
캄브리아기 大爆發
[
編輯
]
古生物學者
들은
化石
을 硏究하던 途中 地球 歷史上 比較的 짧은 期間 內에 動物의 複雜性이 急激하게 增加한 時點이 있음을 밝혀내었다.
[47]
이를
캄브리아기의 大爆發
이라고 부르는데, 이를 둘러싸고 여러 疑問들이 提起되었다. 暴發을 點火시킨 要因은 무엇인가? 캄브리아기의 奇妙한 化石 構造들,
現生 動物
軍들과는 다른 이런 構造들이 現生 動物群들과는 어떠한 關係가 있을까? 이렇게 複雜한 動物들이 何必 이때 처음 登場한 까닭은 무엇인가? 이에 對하여 다양한 假說이 提示되었지만,
發生學
의 劇的인 發展으로 캄브리아기의 大爆發에서 科學者들은 遺傳子가 어떠한 役割을 맡았는지 밝혀낼 수 있게 되었다.
캄브리아기
動物軍들을 보고 第一 먼저 指摘하게 되는 點은 反復되는
身體附屬
의 種類와 數가 다양하게 進化했다는 事實인데, 이는
윌리스턴의 法則
을 劇的으로 드러내는 例이다.
[28]
共通의 툴킷 遺傳子와 스위치의 變化
[
編輯
]
흔히 새로운 種類의 身體 設計와 構造가 登場하기 위해서는 먼저 새로운 遺傳子들이 進化해야 한다고 생각한다. 그러나
節肢動物
의 祖上인
葉族動物
과 가까운
類緣關係
에 있는
有爪動物
과 節肢動物의 號메오 遺傳子를 比較한 結果, 節肢動物의 號메오 遺傳子들이 有爪動物과 節肢動物의 共通 祖上에 이미 存在하고 있었다는 事實이 밝혀졌다.
[48]
[49]
이로써 號메오 遺傳子의 數가 暴發的으로 늘어난 것이
캄브리아기 大爆發
의 原因이라는 假說이 깨어지게 되었고, 科學者들은 이제 새로운 解答을 찾아야만 했다. 이에 對한 代案으로 提示된 것이 스위치의 變化이다. 現生 節肢動物의 設計들을 比較함으로써 科學者들은
號메오 遺傳子
發現 地域이 移動하면서 竊脂 動物 設計에서 主要한 差異들이 생겨났음을 알 수 있었다.
[50]
[51]
[52]
따라서
附屬肢
의 種類 및 機能이 다양해진 것은
體軸
을 따라 늘어선 서로 다른 領域에서 서로 다른 號메오 遺傳子들이 發現했기 때문이라고 할 수 있다. 脊椎 動物의 進化 過程에서도 이와 비슷한 現象이 發見되는데, 初期 脊椎動物의 進化 過程은 캄브리아기 또는 그 以後 生物들의 進化過程과는 달리 號메오 複合體 또는 遺傳體를 複製하여 그 數 自體를 늘였다. 그러나 高等 脊椎動物의 進化 歷史 後半에서는 遺傳子의 數가 重要한 要因이 아니었다. 脊椎動物의 身體 形態 鎭火는 節肢動物과 비슷하게, 體軸에서의 號메오 遺傳子의 發現 地域의 移動으로 이루어졌다.
[53]
[54]
이렇게 胚芽에서
號메오박스
의 座標를 統制하는 것은 바로 號메오 遺傳子의 스위치들이며, 이 스위치들의 DNA 序列이 變化함으로써 벌어지는 일이라는 것이 밝혀짐으로써 드디어 어떻게 動物群이 다양하게 形成되었는지에 對한 실마리가 풀렸다. 萬若
號메오 蛋白質
을 暗號化하고 있는 序列 自體에 變化가 일어났다면 그 모든 機能들에 影響이 미쳤을 것이다. 그러나 實際로는 特定 스위치에만 變化가 일어났기 때문에 다른 身體 附屬들에는 아무런 影響 없이 特定
모듈
萬 變化를 일으킬 수 있었다.
遺傳的 可能性과 生態的 機會의 만남
[
編輯
]
動物의 形態 變化는
툴킷 遺傳子
의 發明 그 自體에 있지 않다는 것이 알려지자, 最近에는
캄브리아기 大爆發
의 原因이
生態的 現象
이라는 意見이 支持를 얻고 있다. 툴킷 遺傳子들은 動物의 形態를 指示해주는 重要한 役割을 하나, 툴킷 自體는 可能性을 의미할 뿐, 運命을 指示하는 것은 아니다. 캄브리아기의 大爆發은 生態的 相互作用과 다양한 動物종이 增加함에 따라 競爭의 壓迫이 갈수록 커져, 보다 複雜한 構造들이 쉴 새 없이 進化한 것으로 봐야 한다.
[55]
遠視 附屬肢의 專門化
[
編輯
]
進化의 歷史는 一種의 ‘軍備 擴張 競爭’이라고 할 수 있다. 같은 環境에서 더 잘 살아남을 수 있게 水泳하고, 걷고, 달리고, 뛰어오르고, 숨 쉬고, 땅을 파고, 날고, 飮食을 잡고, 으깨고, 삼키고, 찌르고, 거르고, 빨아들이는 데 必要한 더 뛰어나고, 빠르고, 가볍고, 剛하고, 敏捷한
附屬肢
들을 가지기 위해 競爭하는 것이었다. 이러한 附屬肢의 進化는 連續的으로 反復되는 身體 設計 德分에 일어날 수 있었다. 두 雙 以上의 構造에 지워져 있던 어떤 機能의 짐을 적은 數의 構造로 옮기고, 그로써 자유롭게 된 構造들을 새 目的에 맞게 專門化시킬 수 있기 때문이다.
[56]
節肢動物의 附屬肢 鎭火
[
編輯
]
節肢動物
의 進化를 論할 때 附屬肢
形態學
은 늘 重要한 主題였다. 그 中心에는 共通 祖上의 이분지(二分枝)型 附屬肢가 있다.
[47]
모든 附屬肢들이
이분지型 附屬肢
에서 出發하여 變形된 것이기 때문이다. 過去의 生物學者들은 外形的 形態學에 全的으로 依存하여 單純한 다리를 가진
昆蟲類
,
지네류
,
노래氣流
,
有鳥類
를 한 動物群으로 묶고, 보다 神奇하게 갈라진 附屬肢를 가진
甲殼類
,
三葉蟲類
,
前葛藟
,
투구게類
를 다른 動物群으로 묶어 두 가지가 다른 種類라고 믿곤 했다.
그러나 갈라진 다리와 갈라지지 않은 다리의 起源이 다를 것이라는 傳統的인 見解와는 달리, 最近의 硏究 結果들은 이분地形 다리는 單純한 管 模樣이었던
葉族動物
의
葉狀族
으로부터 進化한 것 같다는 事實을 支持하고 있다.
[57]
[58]
이를 支持하는 進化發生生物學的 證據도 있다. 有爪動物의 葉狀族과 節肢動物의 다리 盆地에서 共通으로 附屬肢의 形成을 돕는
디스탈리스
遺傳子가 發現되는 것을 確認한 것이다.
[59]
[60]
갈라진 다리나 갈라지지 않은 다리나 모든 節肢動物 附屬肢 形態들은 獨立的으로 發明된 것이 아니라 共通의 古代 葉狀族에서 進化한 것임을 支持하는 事實이다.
날개의 起源
[
編輯
]
昆蟲의 날개의 由來는 오래前부터 論難이 되어온 풀리지 않는 미스터리였다. 날개 없는 昆蟲의
胸部
外皮
가 바깥으로 자라서 날개를 만든 것이라는 主張과 祖上動物의 아가미로부터 날개가 생겨났을 것이라는 主張이 팽팽히 맞서 좀처럼 合意를 이루지 못했다. 이보디보는 이에 對하여 强力하고 確實한 證據를 提示하였다.
美할리스 아베로프
와
스티븐 코언
은 醋파리의 날개 發生 過程을 硏究하여 날개를 形成하는데 必要한 툴킷 蛋白質들을 밝히고, 이 蛋白質들이 다른 節肢動物의 附屬肢에도 發現되는지 調査해보았는데, 놀랍게도
甲殼類
附屬肢의
外分地
, 卽 呼吸 盆地에서 選擇的으로 發現하였다. 이 硏究 結果는 呼吸 盆地와 昆蟲의 날개가
相同器官
이라는 結論을 낳았다. 이는 昆蟲의 날개가 祖上 生物의 아가미에서 나왔다는 理論을 뒷받침하기에 充分했다.
[61]
이보디보의 例示
[
編輯
]
이보디보 硏究의 代表的인 例示로는 나비의 날개무늬와 哺乳類의 黑色症 進化 等이 있다.
나비의 날개
[
編輯
]
그러므로 이 나비들의 널따란 날개膜을 書板 삼아, 自然이 그 위에 鐘의 變形에 對한 이야기를 써둔 것이라고 할 수 있다. 實로 自然 組織의 모든 變化들이 그 위에 간직되어 있다고도 볼 수 있을 것이다. […] 게다가 날개의 色깔 패턴은 一般的으로 種間의 血緣 程度를 말해주는 썩 規則的인 證據가 된다. 自然의 法則은 모든 存在들에게 同一할 것이므로 昆蟲의 한 種類인 이들에게서 끌어낸 結論은 全體 有機體 世界에 適用되어야만 한다. 따라서 나비를 硏究한다는 것은, 設令 산뜻하고 하늘하늘한 生命體라서 選擇한 對象이라 하더라도, 輕蔑 받을 일이 아니다. 도리어 언젠가 生物科學의 가지들 中에서도 가장 重要한 硏究로 인정받을 것이다.
英國의 박물학자
헨리 월터 베이츠
가 그의 冊 『아마존 講義 박물학자』에서 나비를 言及한 뒤로, 나비는 進化的으로, 또 生態學的으로 흥미로운 主題가 되었다.
[62]
그 中에서도 이보디보는 나비가 어떻게 그렇게 다양하고 아름다운 무늬를 가질 수 있게 되었는지에 對해 說明한다. 흔히 나비의 날개에는 어떠한 規則性도 없다고 생각하지만, 1920年代와 1930年代에 몇몇
比較生物學
者들이 가장 理想的인 狀態의 나비 무늬의 設計圖를 作成하였다.
[63]
各各의 나비 종들은 이 基本 設計 計劃으로부터 다양한 水準으로 벗어난
變異
라고 말할 수 있다. 現實의 나비 날개 무늬들은 各各의 무늬 要素들을 最大로 包含하는 基本 設計와는 달리 特定 要素들이 强調되거나 省略되고 조금씩 變形되고는 한다. 이러한 나비의 날개를 觀察함으로써 알 수 있는 事實은 서로 다른 무늬 要素들이 다른 무늬 要素들과는 相關없이 獨自的으로 形態와 色과 크기를 바꾸는 方向으로 進化할 수 있다는 點이다. 脊椎動物이나 節肢動物에서 본
모듈
構造가 나비의 날개救助 鎭火에서도 나타나고 있다는 事實을 알 수 있다.
 나비의 눈꼴무늬
나비의 눈꼴무늬
多樣한 나비 날개무늬의 構成 要素 中에서도 科學者들이 焦點을 맞춘 것은 나비의
눈꼴무늬
에 對해서였다. 눈꼴무늬는 點박이 形態로, 色이 다른
人便
들이 여러 겹의 同心圓을 이루어 만들어진다. 눈꼴무늬는
捕食者
의 視線을 相對的으로 軟弱한 몸통으로부터 分散시켜 날개 가장자리로 向하게 함으로써 捕食者의 襲擊으로부터 나비를 防禦하는데 核心的인 役割을 맡는다.
1980年
듀트 大學
의
프레드 네以下우트
는 未來의 눈꼴무늬 位置가
幼蟲
段階에서 이미 決定된다는 事實을 밝혀냈다.
[63]
그 後
션 캐럴
과 그의 硏究팀은 醋파리의 날개를 形成하는
號메오 遺傳子
가 나비에도 存在한다는 것을 確認하고, 그 中에서도 눈꼴무늬를 形成하는데 關與하는 遺傳子는 醋파리 및 節肢動物의 附屬肢 形成에 關與하는 遺傳子인
디스탈리스
라는 것을 알아냈다.
[64]
[65]
얼핏 이 事實을 디스탈리스가 醋파리와 나비에서 各各 다른 役割을 한다는 것을 의미한다는 것으로 誤解하기 쉬우나, 곧 디스탈리스가 나비에서도 附屬肢 形成에 如前히 關與하고 있다는 事實이 드러났다. 디스탈리스는 特定 位置 및 時期에 變함없이 四肢를 形成하는 役割을 遂行한다. 다만 날개 무늬에서의 作業은 그와는 또 다른 位置 및 時期의 일로서, 全혀 다른 形態로 統制되는 나비만의 무늬 形成 스위치를 進化시킨 것에 對한 結果라고 볼 수 있다. 卽, 本來의 遺傳子에 새로운 스위치들이 鎭火함으로써 나비에서 새로운 任務를 遂行할 수 있게 된 것이다.
哺乳類의 黑色症 鎭火
[
編輯
]
動物의 身體 附屬의 一部나 몸 全體가 어둡게 着色되는 現象은 自然에서 가장 흔하게 登場하는 進化的 變化이다. 이에 對한 硏究는 오래前부터 進行되어 왔고, 이러한 色素 變化에 어떤
選擇압
이 作用했는지,
形質
進化의
分子生物學
敵 起源은 무엇인지 밝혀진 事例들도 있다. 이보디보의 發電으로 硏究者들은 哺乳類의 色, 特히 검은色의 進化에 對하여 더 많은 것을 밝힐 수 있었다.
遺傳子가 어떻게 動物의 色素 形成에 影響을 끼치는지 알아볼 수 있는 가장 두드러진 現象으로는
黑色症
이 있다. 黑色症이란 어떤 個體나 鐘이 元來의 色이 있어야 하는 자리가 代身 검은 系統의 色깔로 着色되는 現象을 말한다. 黑色症의 原因은
멜라닌 色素
로써, 다양한 形態와 黃褐色부터 검은色에 이르기까지 넓은 範圍의 어두운 色을 띤다. 黑色症의 起源에 對해 紫外線 損傷으로부터의 保護, 體溫 調節,
擬態
, 或은 異性에게 選擇 받는 데(
星間選擇
)에 유리하다는 여러 說들이 있지만, 正確한 理由는 밝혀지지 않았다.
[66]
黑色症과
自然選擇
에 對한 有名한 事例로
産業革命
以來로 變化한 英國 및 美國 北部의 産業地球에 사는
얼룩나방
의 分布가 있다. 얼룩나방은
티피카
形態와
카르보나리兒
形態의 두 가지가 存在하는데, 티피카 形態는 흰 바탕에 검은 얼룩이 나 있고, 카르보나리兒 形態는 全體가 다 새카맣다. 産業化가 일어난 地域의 나무들은 그을려서 劍氣 때문에 티避카 形態보다 카르보나리兒 形態의 얼룩나방이 몸을 숨기기에 有利했던 反面, 티피카 形態는 밝은 곳에,
이끼
가 나 있는 나무에서 더 살아남기 유리했다.
[67]
[68]
[69]
또한
재규어
와 같은
큰 고양이과
動物의 黑色症도 잘 알려져 있는 例 中 하나이다.
[70]
哺乳類
는 皮膚 및 毛囊의
色素 細胞
에서 두 가지의
멜라닌 色素
를 만든다.
有멜라닌
은 털에 黑褐色을 입히고,
페오멜라닌
은 丹紅色 또는 노란色을 입힌다. 멜라닌 色素의 量은 여러 蛋白質들에 依해 調節되는데, 特히
멜라盧코르틴-1 受容體(MC1R)
蛋白質이 重要하게 作用한다. MC1R 受容體는 細胞膜을 사이에 두고 細胞 안팎으로 뻗어있는데,
알파-멜라닌 細胞 刺戟 호르몬(MSH)
과 結合하면 色素 細胞로 有멜라닌을 合成하라는 信號가 傳達한다. 反對로,
아구티
라는 蛋白質이 이 受容體에 結合해서 抑制字로 作用하면 페오멜라닌이 生成된다. 正常的인 黃色 재규어와 黑色症 재규어를 比較해보니, 黑色症 재규어의 MC1R 遺傳子에 생긴
優性 突然變異
로 因해 MC1R 蛋白質의
아미노산
序列에 問題가 생겨 黑褐色을 만드는 有멜라닌 合成이 持續되었음을 알 수 있었다. MC1R 受容體가 聯關된
호르몬族
들과 密接하게 反應함에도 不拘하고, 受容體의 變化가 일어날 수 있었던 것은 이것이 大體로 色素 調節에만 使用되기 때문이었다.
[71]
MC1R 突然變異는 재규어 以外의 종들에서도 黑色症의 原因이 된다.
[72]
[73]
[74]
놀라운 點은, 이렇게 MC1R 遺傳子에 突然變異가 생겨나서 黑色症이 생기는 鐘들이 만드는 아미노산이 모두 同一하다는 것이다. 다시 말해, 野生 고양이類와 潮流가 同一한 蛋白質에 獨立的으로 突然變異를 일으켜 비슷한 進化的 變化를 낳았다는 것이다. 遺傳子의 水準뿐만 아니라 蛋白質의 아미노산 水準에서까지 進化가 反復될 수 있음을 決定的으로 보여준 것이었다.
實際로 自然에서는 몸 全體에 한 色이 쓰인 境遇보다는 主로 둘 以上의 色들이 空間的으로 複雜하게 分布한 境遇가 더 많다. 이것은 發生에서 遺傳子의 調節이
모듈
單位로 일어나기 때문에 可能한 일이다. 遺傳子가 한 場所에서는 選擇的으로 發現하고 다른 場所에서는 發現하지 않으려면, 色素 遺傳子들의 發現 및 色깔 패턴을 統制하는 스위치들이 있어야 한다.
哺乳類의 무늬에 關한 硏究 中에서 가장 흥미로우면서 가장 밝혀지지 않은 問題는 아마도
얼룩말
의 무늬에 關한 것일 것이다.
[75]
그 中에서도
조너선 바드
는 現生 얼룩말 종들 사이에서 줄무늬 個數가 差異 나는 것에 關心을 가졌다. 바드는 各 種의 發生 過程에서 멜라닌 細胞의 移動 始作 時點이 다르기 때문에 줄무늬 個數가 다른 것이라고 主張했다. 바드가 注目한 點은 줄무늬 個數가 적으면 무늬 幅이 넓고, 個數가 많으면 무늬 幅이 좁다는 事實이다. 바드는 모든 初期 胚芽에서 줄무늬들이 發生하는 間隔은 일정하지만, 種마다 發生 始作 時點이 다르기 때문에 생기는 現象이라고 推測했다. 무늬 發生이 일찍 始作될수록 무늬 幅은 넓어지고 몸에 그려질 무늬의 個數는 줄어들 것이다. 거꾸로 무늬 發生이 늦게 始作되면 全體 胚芽의 크기에 比해 무늬 幅이 좁게 될 것이고, 當然히 더 많은 數가 그려질 것이다.
[76]
같이 보기
[
編輯
]
參考 文獻
[
編輯
]
- 『이보디보(生命의 블랙박스를 열다)』 션 B. 캐럴 저/ 김명남 驛/ 지호/ 2007.07.23
- 『내안의 물고기(물고기에서 人間까지, 35億 年 進化의 祕密』 닐 슈빈 저/ 김명남 驛/ 김영사/ 2009.06.08
- 『한치의 疑心도 없는 鎭火 이야기(DNA와 進化의 確固한 證據들)』 션 B. 캐럴 저/ 김명주 驛/ 지호/ 2008.10.13
- 『發生學 길라잡이』 스콧 F. 길버트 저/ 剛해묵 驛/ 라이프사이언스/ 2011.09.01
- 김창배 (韓國生命工學硏究院),
鎭火發生生物學
, 生物學專門硏究센터, 2003 4月號</ref>
읽어 보기
[
編輯
]
- 『이보디보(生命의 블랙박스를 열다)』 션 B. 캐럴 저/ 김명남 驛/ 지호/ 2007.07.23
- 『내안의 물고기(물고기에서 人間까지, 35億 年 進化의 祕密)』 닐 슈빈 저/ 김명남 驛/ 김영사/ 2009.06.08
- 『한치의 疑心도 없는 鎭火 이야기(DNA와 進化의 確固한 證據들)』 션 B. 캐럴 저/ 김명주 驛/ 지호/ 2008.10.13
- EvoDevo 情報의 鎭火
, 한겨레新聞, 2008/05/02
各州
[
編輯
]
- ↑
가
나
김창배(상명대), 遺傳子에서 찾는 21世紀版 進化法則, 科學東亞 2009年 1月號, 동아사이언스, ISBN AB-D2-00901-0
- ↑
가
나
김창배, 鎭火發生生物學, (2003) - 參考 文獻을 參照할 것
- ↑
가
나
최재천(서울大), 分子生物學 萬能 時代 克服하는 統合 生物學 - 進化와 發生으로 아우르는 이보-디보의 첫걸음, 科學東亞 2003年 4月號, 동아사이언스, ISBN AB-D2-00304-0
- ↑
Tomarev, Stanislav I.; Callaerts, Patrick; Kos, Lidia; Zinovieva, Rina; Halder, Georg; Gehring, Walter; Piatigorsky, Joram (1997).
“Squid Pax-6 and eye development”
. 《Proceedings of the National Academy of Sciences》
94
(6): 2421?2426.
Bibcode
:
1997PNAS...94.2421T
.
doi
:
10.1073/pnas.94.6.2421
.
PMC
20103
.
PMID
9122210
.
- ↑
Pichaud, Franck; Desplan, Claude (2002年 8月). “Pax genes and eye organogenesis”. 《Current opinion in genetics and development》
12
(4): 430?434.
doi
:
10.1016/S0959-437X(02)00321-0
.
PMID
12100888
.
- ↑
Pennisi, E (2002). “EVOLUTIONARY BIOLOGY:Evo-Devo Enthusiasts Get Down to Details”. 《Science》
298
(5595): 953?955.
doi
:
10.1126/science.298.5595.953
.
PMID
12411686
.
- ↑
션 캐럴. 《이보디보 : 生命의 블랙박스를 열다》. 지호 ,
ISBN
978-89-8909-029-7
{{
isbn
}}의 變數 誤謬: 有效하지 않은
ISBN
.
.
- ↑
Bowler 2003
, 120~128, 208쪽
括弧 없는 하버드 認容 error: 對象 없음: CITEREFBowler2003 (
help
)
Secord 2003
, 424, 512쪽
括弧 없는 하버드 認容 error: 對象 없음: CITEREFSecord2003 (
help
)
- ↑
Desmond & Moore 1991
, 490?491쪽
括弧 없는 하버드 認容 error: 對象 없음: CITEREFDesmondMoore1991 (
help
)
- ↑
Darwin, Charles (1859). On the Origin of Species (1st ed.). London: John Murray. p. 1.
ISBN
0-8014-1319-2
.
- ↑
Bowler 2003
, 170, 190~191쪽
括弧 없는 하버드 認容 error: 對象 없음: CITEREFBowler2003 (
help
)
- ↑
Ridley, Mark (2003).
《Evolution》
. Wiley-Blackwell.
ISBN
978-1-4051-0345-9
.
- ↑
Gould, Stephen Jay
(1977). 《
Ontogeny and Phylogeny
》. Cambridge, Massachusetts: Harvard University Press.
ISBN
0-674-63940-5
.
- ↑
高級生命科學, 이길재, 서울特別市敎育廳
- ↑
[1]
- ↑
Scott MP, Tamkun JW, Hartzell III GW (1989). “The structure and function of the homeodomain”. 《Biochim. Biophys. Acta》
989
(1): 25?48.
PMID
2568852
.
- ↑
Gehring WJ (1992). “The homeobox in perspective”. 《Trends Biochem. Sci.》
17
(8): 277?280.
doi
:
10.1016/0968-0004(92)90434-B
.
PMID
1357790
.
- ↑
가
나
Schofield PN (1987). “Patterns, puzzles and paradigms - The riddle of the homeobox”. 《Trends Neurosci.》
10
: 3?6.
doi
:
10.1016/0166-2236(87)90113-5
.
- ↑
Alonso CR (2002). “Hox proteins: sculpting body parts by activating localized cell death”. 《Curr. Biol.》
12
(22): ?.
doi
:
10.1016/S0960-9822(02)01291-5
.
PMID
12445403
.
- ↑
Carrasco, McGinnis, Gehring and De Robertis, Cell 37, 409-414, 1984
- ↑
Ryan, Joseph F; Maureen E. Mazza, Kevin Pang, David Q. Matus, Andreas D. Baxevanis, Mark Q. Martindale, John R. Finnertyl (2007年 1月). Fay, Justin, 編輯.
“Pre-Bilaterian Origins of the Hox Cluster and the Hox Code: Evidence from the Sea Anemone, Nematostella vectensis”
. 《PLoS ONE》
2
(1): e153.
doi
:
10.1371/journal.pone.0000153
.
PMC
1779807
.
PMID
17252055
. 2008年 4月 30日에 確認함
.
- ↑
R.Riddle et al. 『cell』 75(1993), pp. 1401~1416
- ↑
가
나
『Nature』, 287(1980), pp 795~801
- ↑
W. McFinnis et al, 『Cell』 37(1984), pp.403~408
- ↑
D. Douboule and P. Dolle, 『EMBO Journal』 8 (1989), pp. 1497~1505
- ↑
A. Graham, N. Papalapov, and R. Krumlauf, 『Cell』 67(1989) pp.367~378
- ↑
R. Quiring et al, 『Science』 265(1994), pp.785~789
- ↑
가
나
Williston, Samuel Wendall (1914). 『Water Reptiles of the Past and Present』 Chicago: University of Chicago Press.
- ↑
H. Spemann, 『Embryonic Development and Induction』 (New Haven: Yale University Press, 1938)
- ↑
R. Fleischmajer and R.E.Bilingham eds. 『Epithelial Mesenchymal Interactions』 (Baltimore: Williams and Wilkins, 1968)
- ↑
Allen, Garfield E.
Thomas Hunt Morgan: the man and his science
. Princeton University Press 1978
- ↑
E.Lewis, 『Nature』 276 (1978) pp. 565~570
- ↑
B. Wakimoto and T.Kaufamn, 『Developmental Biology』 81 (1981), pp.51~64
- ↑
A. S. Laughon and M. P. Scott, 『Nature』 310 (1984) pp.25~31
- ↑
Peter Lawrence 『The Making of the Fly』 (Medford, Mass. : Blackwell Science, 1922)
- ↑
W. Gehring 『Master control genes in the development and Evolution: The Homeobox Story』 (New Haven : Yale University Press, 1999)
- ↑
가
나
McGinnis, William; Michael Kuziora (1994年 2月). “The Molecular Architects of Body Design”. 《
Scientific American
》
270
(2): 36?42.
PMID
7906436
.
- ↑
가
나
D.Duboule and P. Dolle 『EMBO Journal』 8(1989), pp. 1497~1505
- ↑
A. Graham, N. Papalapov, N. Papalapov, and R.Krumlauf, 『Cell』 57 (1989) pp. 367~378
- ↑
R.Quiting et al, 『Science』 265 (1994) pp. 785~789
- ↑
가
나
G. Halder, P. Callaerts, and W. Gehring 『Science』 267 (1994) pp. 1788~1792
- ↑
가
나
G. Panganiban et al. 『Proceedings of the National Academy of Science, USA』 94 (1997) pp. 5162 ~ 5166
- ↑
R. Bodmer and T.V. Venkatregh, 『Developmental Genetics』 22(1998) pp. 181~186
- ↑
W.Bateson 『Materials for the Study of Variation』 (London : Macmilan, 1894)
- ↑
션 캐럴 『이보디보 : 生命의 블랙박스를 열다』 p.116~117, 지호,
ISBN
978-89-8909-029-7
{{
isbn
}}의 變數 誤謬: 有效하지 않은
ISBN
.
- ↑
션 캐럴 『이보디보 : 生命의 블랙박스를 열다』 p.251~252, 지호,
ISBN
978-89-8909-029-7
{{
isbn
}}의 變數 誤謬: 有效하지 않은
ISBN
.
- ↑
가
나
S.J.Gould 『Wonderful Life: The Burgess Shale and the Nature of History』 (New York : W. W. Norton, 1989)
- ↑
E.B.Lewis 『Nature』 276(1978) pp.565~570
- ↑
J. K. Grenier et al 『Current Biology』 7 (1997) pp.547~553
- ↑
M.Averof and M. Akam 『Nature』 376(1995) pp. 547~553
- ↑
M. Averof and N.H. Patel 『Nature』 338 (1997) pp. 682~687
- ↑
S. B. Carroll 『Nature』 376(1995) pp.479~485
- ↑
S.M. Shimeld and P. W. Holland 『Proceeding of the National Academy of Science, USA』 97 pp.4449~4452; (2000)
- ↑
H. G. Belting, C. Shashikant, and F. H. Ruddle 『Proceedings of the National Academy of Science, USA』 95 (1998) pp. 2355~2360
- ↑
Andrew H. Knoll 『Life on a Young Planet : The First Three Billion Years of Evolution on Earth』 (Princeton : Princeton University Press, 2003)
- ↑
션 캐럴 『이보디보 : 生命의 블랙박스를 열다』 p.221, 지호,
ISBN
978-89-8909-029-7
{{
isbn
}}의 變數 誤謬: 有效하지 않은
ISBN
.
- ↑
G. E. Budd 『Lethaia』 29 (1996) pp.1~14
- ↑
N. Shubin, C. Tabin, and S. Carroll, 『Nature』 388(1997) pp. 639~648
- ↑
G. Panganiban et al 『Science』 270(1995) pp.1363~1366
- ↑
Panganiban et al 『Proceedings sof the National Academy of Sciences, USA』 94 (1997) pp. 5162~5166
- ↑
M. Averof and S. M. Cohen 『Nature』 385 (1997) pp. 627~630
- ↑
Henry Walter Bates 『Naturalist on the River Amazons』 (London: John Murray, 1863)
- ↑
가
나
Frederick Jihout 『The Development and Evolution of Butterfly Wind Patterns』 (Washington D.C. : Smithsonian Instituion Press, 1991)
- ↑
S. B. Carroll et al. 『Science』 265(1994) pp.109~114
- ↑
S. B. Carroll 『Natural History』 February 1997 pp.28~37
- ↑
M. Majerus 『Melanism : Evolution in Action』(Oxford: Oxford University Press, 1988)
- ↑
B. N. Grant 『Evolution』 53(1999) pp. 980~984
- ↑
J. Mallet 『Genetics Society News』 50 (2003) pp.34~38
- ↑
J. Hopper 『Of Moths and Men : Intrugue, Tragedyand the Peppered Moth』 (New York: Fourth Estate, 2002)
- ↑
M. E. N. Majerus and N. I. Mundy 『Trends in Genetics』 19(2003) pp. 585~588
- ↑
E. Eizirik et al 『Current Biology』 13(2003) pp.448~453
- ↑
E. Theron et al 『Current Biology』 11(2011) pp. 550~557
- ↑
M. Nachman et al 『Proceedings of the National Academy of Science, USA』 100 (2003) pp. 5268~5273
- ↑
K. Ritland et al 『Current Biology』 11(2011) pp. 1468~1472
- ↑
S. J. Gould 『Hen’s Teeth and Horse’s Toes』 (New Tork: W. W. Norton, 1983) pp. 355~365, pp. 366~375
- ↑
J. L. Bard 『Journal of Zoology』(London) 183 (1997) pp.527~539