吐露社友루스
(
Torosaurus
, 구멍이 있는 도마뱀이란 뜻으로, 프릴에 있는 큰 구멍을 가리키는 이름)는
白堊紀
後記
마스트리흐트節
後記 約 6800萬年에서 6600萬年 前에 살았던 草食性
케라톱스과
恐龍
의 한
속
이다. 化石은 北아메리카 西部內陸, 캐나다의
사스캐처원
州에서
텍사스
南部에 이르는 地域에서 發見된다.
吐露社友루스의 頭蓋骨은 알려진 陸上動物들의 頭蓋骨 中 가장 큰 축에 屬한다. 프릴이 달린 頭蓋骨의 길이는 2.77m 에 達한다.
트리케라톱스
와 비슷한 크기였을 것으로 推定된다.
[1]
[2]
[3]
몸무게는 6톤 程度로 推定된다. 吐露社友루스는 긴 프릴에 커다란 구멍들이 있고 프릴의 一部인 비늘뼈의 위쪽 表面에 고랑이 파여 있으며 프릴 뒤쪽에 다섯 雙 以上의 작은 뿔 (뒷머리뼈突起) 들이 있어 同時代에 살았던 트리케라톱스와는 區別된다.
[4]
吐露社友루스는 또 트리케라톱스 프로르수스에서 볼 수 있는 긴 코뿔이 아니라 代身 더 이른 時期의 相對的으로 原始的인 트리케라톱스 호리두스와 類似한 짧은 코뿔을 가지고 있다.
[4]
吐露社友루스 라투스, 吐露社友루스 글라디우스, 吐露社友루스 우타헨시스 세 種이 命名되었다. 하지만
T. gladius
는 有效한 種으로 看做되지 않는다.
最近 吐露社友루스의 有效性이 論爭의 對象이 되었다.
[5]
2010年 化石 骨
組織學
과 프릴의 模樣을 調査한 硏究에서 吐露사우루스가 아마도 트리케라톱스의 聖體 形態인 것 같다는 結論을 내리고 있다. 典型的인 트리케라톱스 標本의 뼈들이 未成熟한 狀態이며 吐露社友루스의 特徵인 프릴의 구멍이 생겨나기 始作하는 兆朕을 보인다는 것이다. 個體가 成熟하는 過程에서 頭蓋骨의 프릴은 엄청나게 길어지며 구멍이 나타날 수 있다는 主張이다.
[6]
[7]
[8]
하지만 2011, 2012, 그리고 2013年에 各各 알려진 標本들의 特徵들을 硏究하여 두 속의 形態的인 差異를 보면 이들이
動物耳鳴
일 可能性은 없다고 主張하는 硏究가 出版되었다. 吐露사우루스가 트리케라톱스의 聖體라는 主張의 주된 問題는 分明한 中間 形態가 없다는 點이다. 吐露사우루스가 분명하지만 頭蓋骨의 길이 比率이 다른 아성체가 存在하고, 成體가 되면서 프릴에 구멍이 생기는 것이 각룡류의 一般的인 成熟 過程이 아니라는 것이다.
[4]
[9]
[10]
發見과 種
[
編輯
]
트리케라톱스
가 命名된 지 2年 後인 1891年,
와이오밍
州 南東部 니오브라라 카운티에서 존 벨 해처가 긴 프릴에 구멍이 있는 한 雙의 角龍類 頭蓋骨을 發見했다. 해처를 雇用했던 古生物學者
오트니엘 찰스 마쉬
가 吐露社友루스라는 俗名을 지었다.
[11]
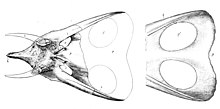 T. latus
와 그 이명인
T. gladius
頭蓋骨을 그린 마쉬의 원 圖解
T. latus
와 그 이명인
T. gladius
頭蓋骨을 그린 마쉬의 원 圖解
吐露社友루스라는 이름은 種種
라틴語
名士
taurus
, 或은 스페인語 "吐露" 로 誤認되어 "황소 도마뱀" 이라고 불리곤 하는데,
古代그리스어
로 "구멍을 뚫다"라는 意味의 動詞인 "토레誤"
τορ?ω
에서 由來했을 可能性이 훨씬 높다.
[12]
이것은 吐露社友루스의 이름이 길쭉한 프릴에 있는 구멍을 가리킨다고 보는 것인데, 트리케라톱스의 프릴에는 구멍이 없기 때문에 吐露社友루스와 트리케라톱스를 區分하는 方法으로 吐露社友루스 프릴의 구멍이 使用되어 왔다. 吐露社友루스의 이름에 混同이 있는 것은 마쉬가 元來의 論文에서 이름의 由來를 說明하지 않았기 때문이다.
吐露社友루스에는 두 種이 存在한다.
- T. latus
Marsh, 1891
(
모식종
). 라투스는 "넓다"는 意味로 프릴의 形態를 가리킨다.
- T. utahensis
Lawson, 1976
.
後에 命名된
T. gladius
種은
T. latus
의 異名으로 看做된다.
- T. gladius
Marsh, 1891
. 글라디우스는 라틴語로 "칼"을 意味한다. 비늘뼈의 긴 形態를 가리킨다.
T. latus
의
완모式標本
은
YPM 1830
으로 頭蓋骨 一部로 構成되어 있다.
T. gladius
의 완모式標本은 YPM 1831 로 더 큰 頭蓋骨이다. 두 標本 모두
마스트리흐트節
의 랜스 層에서 發見되었다. 그 以後 비슷한 標本들이
와이오밍
,
몬태나
,
사우스다코타
,
노스다코타
,
콜로라도
,
유타
, 그리고
사스캐처원
州 等에서 發見되어 吐露社友루스로 分類되었다. 確實하게 分類가 可能한 標本들로는 다음과 같은 것들이 있다. ANSP 15192는 에드윈 해리스 콜버트가 1944年에
사우스다코타
에서 發見한 작은 個體이다.
[13]
MPM VP6841 은 頭蓋骨 一部로 現在는
밀워키
에 展示되어 있다. SMM P97.6.1 은 주둥이가 없는 頭蓋骨이다. 헬크릭 層에서 發見된 두 個의 頭蓋骨 一部는 2002年에 報告되었고, MOR 981 은 1998年에 MOR 1122는 2001年에 發見되었다.
[14]
吐露社友루스라고 分類될 可能性이 있는 斷片的인 化石들이
텍사스
의 빅벤드 地域과 뉴멕시코의 산후안 盆地에서 發見되었다.
[15]
吐露社友루스 標本은 相對的으로 드물고 트리케라톱스가 훨씬 豐富한 便이라고 한다.
吐露社友루스 우타헨시스 (
Torosaurus utahensis
)는 찰스 휘트니 길모어가 1946年에 元來
아르히노케라톱스 우타헨시스
로 報告했고, 標本인 USNM 15583는
유타
注意 에머리 카운티에서 發見된 프릴 조각으로 構成되어 있다.
[16]
1976年에 더글라스 로슨이 이것을 吐露社友루스 右打헨시스로 재명명하였다.
[17]
로버트 설리반과 同僚들의 2005年 硏究에서는 이것을 吐露社友루스 右打헨시스로 보았고,
T. latus
보다 조금 더 오래된 것으로 看做하였다.
[18]
2008年에 레베카 헌트가 이 種으로 分類될 수 있는 追加 標本을 相當量 報告하였다.
[19]
하지만 後續 硏究에서 이 標本들이 아르히노케라톱스이거나, 或은 새로운 속일 可能性이 提起되었다. 北쪽 헬크릭 層의 恐龍들과 南쪽 "
알라모社友루스
動物群"은 서로 겹치는 境遇가 잘 없으며 아마도 地理的 障壁으로 隔離되어 있었던 것으로 보이기 때문이다.
T. utahensis
가 새로운 속으로 看做되어야 할지, 或은
T. latus
처럼 어떤 트리케라톱스 鐘의 다 자란 成長段階인지에 對한 硏究는 아직 出版되지 않았다.
[6]
特徵
[
編輯
]
 T. latus
復元도
T. latus
復元도
吐露社友루스로 分類되는 個體들은 모두 매우 커서 가장 큰 트리케라톱스 標本과 比較될 수 있을 程度다. 길쭉한 프릴 때문에 特히 頭蓋骨이 긴 便이다. 해처는 YPM 1830 의 頭蓋骨 길이를 2.2m, YPM 1831 을 2.35m 로 推定했다.
[20]
1933年에 리차드 스완 럴은 이것을 各各 2.4m 와 2.57m 로 上向調整했다.
[21]
이 數値로 보자면 吐露社友루스는 陸上 動物 中 가장 긴 頭蓋骨을 가진 것으로 看做된다. 1998年에 토마스 레만은 一部만 남아 있는
펜타케라톱스
의 頭蓋骨 標本 全體 길이가 2.9m 에 達할 것이라고 主張했다.
[22]
이 主張은 2011年에 니콜라스 롱리치는 이 主張에 疑懼心을 表示하며 該當 標本이 너무 길게 復元되었다고 結論내리고 別途의 속인
티타노케라톱스
로 再分類하였다.
[23]
2006年에 앤드류 派키는 해처의 元來 標本들보다 더 긴 두 個의 새로운 頭蓋骨을 報告하였다. MOR 1122 의 길이는 252 cm, MOR 981 은 277 cm이다.
[24]
 MOR 1122 標本
MOR 1122 標本
派키는 2006年에 吐露社友루스의 分類可能한 特徵을 確立했다. 프릴은 頭蓋骨의 다른 部分에 비해 極端的으로 길다. 프릴의 뒤쪽, 마루뼈 가장자리에는 三角形 模樣의 骨片人 마루뼈뒷突起가 열 個 以上 있다. 中心線에는 마루뼈뒷突起는 없다. 마루뼈-비늘뼈 境界에 걸쳐 있는 骨片 亦是 없다.
마루뼈
는 얇다. 마루뼈에는 둥글거나 楕圓形의 구멍이 있다. 마루뼈의 너비는 길이보다 20% 程度 넓다. 派키는
T. latus
가 트리케라톱스 호리두스와
T. utahensis
와 區分되는 特徵으로 비늘뼈와 마루뼈가 만나는 部分의 가장자리가 길게 突出되어 있으며 그와 平行하게 골이 파져 있다는 點을 들었다.
[24]
派키는 吐露社友루스의 標本들이 相當한 變異를 보인다고 指摘했다. 눈 위의 뿔들은 때로는 MOR 981처럼 크고 앞으로 휘어져 있고 때로는 MOR 1122 와 ANSP 15191처럼 짧고 똑바르다. 또 이 뿔들의 位置도 差異가 있어서 많은 境遇 눈 바로 위에 位置하지만 YPM 1831 같은 境遇 눈확의 뒤쪽 가장자리에 뿔이 位置한다. 코뿔의 境遇에도 形態에 變異를 보이는데, 곧게 뻗은 코뿔을 가진 YPM 1831 과 그보다 조금 덜하게 곧은 YPM 1830 이 있는가 하면 MOR 981, ANSP 15192, 그리고 特히 MOR 1122는 뿔이라기보다 낮은 突出部를 가지고 있다. 프릴度 亦是 形態에 差異가 있는데 ANSP 15192 와 YPM 1830 은 뒤쪽이 위로 휘어 있는 防牌 模樣이나 YPM 1831는 거의 平平한 模樣이다. 하지만 이것은 復元 過程에서 變形되었을 可能性도 있다. YPM 1831 의 프릴은 하트 模樣으로 中心線 뒤쪽이 움푹 들어가 있으나 다른 標本들은 뒤쪽 가장자리가 곧은 便이다. 프릴의 比率도 相當히 便히가 많다. YPM 1831 의 길이-너비 比率은 1.26 이나 MOR 981 은 길이가 너비의 2.28倍이다. 마루뼈뒷突起의 數는 大槪의 標本들에서 發見되지 않기 때문에 正確히 알기 힘들다. MOR 981 과 MOR 1122는 各各 열 個와 열두 個의 마루뼈뒷突起를 가지고 있다. YPM 1831 은 頭蓋骨 위쪽에 숨구멍이 있는 形態로 復元되었는데, 아마 實際로도 있었을 可能性이 있다. 派키는 吐露社友루스에서 볼 수 있는 變異의 程度가 關聯된 속들에서 볼 수 있는 것보다 크지 않다고 結論내렸다.
[24]
派키는 프릴을 除外하면 吐露社友루스와 트리케라톱스 사이에 특별한 差異는 보이지 않는다는 點을 强調했다. 吐露社友루스 標本은 모두 긴 코뿔을 가지고 있지 않으며 뿔 앞쪽 基底部에 水平으로 動脈이 지나가는 홈도 없지만 트리케라톱스의 境遇도 이러한 特徵들을 보여주는 化石들이 드물지 않다.
[24]
헌트는 2008年에
T. utahensis
가
T. latus
와는 달리 中心線에 마루뼈뒷突起를 가지고 있다는 點에서 트리케라톱스와 類似하다는 結論을 내렸다.
[19]
分類
[
編輯
]
1891年에 마쉬는 吐露사우루스를
角龍類
內의
케라톱스과
로 分類했다.
[11]
각룡류는
鸚鵡새
같은 부리를 가지고
쥐라紀
와
白堊紀
에
北아메리카
와
아시아
에서 蕃盛했던 草食性 恐龍 無理다.
긴 프릴을 가진 吐露社友루스는 傳統的으로
카스모社友루스亞科
로 分類되어 왔다. 吐露社友루스는
안키케라톱스
或은
아르히노케라톱스
의 後孫들로 이루어진 系統에서 뒤늦게 나타난 一員으로 看做되었다. 따라서 1980年代에 들어서 짧은 프릴 때문에
켄트로사우루스아과
로 分類되어온
트리케라톱스
와는 다른 系統으로 생각되었다. 하지만 1990年代에
盆地學
敵 分析에서 이 둘이 모두 카스모社友루스亞科에 屬하는 것으로 나타났다. 最近의 分析들은 모두 吐露社友루스와
트리케라톱스
가 가까운 關係라는 것을 보여주고 있다.
[24]
[25]
트리케라톱스와 같은 種類認知 아닌지에 對한 論爭
[
編輯
]
마스트리흐트節
의
라라미디아
大陸에는 同一한
棲息地
에 살고 있으며 서로 가까운 關係인 카스모社友루스아과의 속이 둘 있었다. 이 둘 사이의 눈에 띄는 差異는 프릴의 形態뿐이었다. 어린 吐露社友루스는 發見된 적이 없으며 어린 트리케라톱스는 相當數가 發見되었다. 트리케라톱스는 다른 카스모社友루스亞科 恐龍들과는 달리 成體가 되어서도 어릴 때의 特徵, 卽 비늘뼈가 짧다는 特徵을 維持하고 있어서
幼形進化
(paedomorphosis)의 例를 보여준다. 2009年에 존 스카넬라는
몬태나
注意 헬크릭 層에서 恐龍의
個體發生
을 硏究하면서 트리케라톱스와 吐露사우루스가 한 속의 서로 다른 成長段階를 나타내는 것이라는 假說이 이런 現象을 가장 잘 說明할 수 있다고 結論내렸다. 吐露社友루스 標本들은 完全히 成長한 트리케라톱스 個體라는 것이다. 이 境遇 吐露社友루스는 트리케라톱스의 動物耳鳴이 되며 트리케라톱스가 優先權을 가지게 된다.
[26]
 "吐露型" 假說에 따르면 트리케라톱스 牙城體 (A, 트리케라톱스 프로르수스 완모式標本 YPM 1822)의 프릴이 길어지고 구멍이 생겨 B, 吐露社友루스 라투스 標本 ANSP 15192 가 되었을 것이다
"吐露型" 假說에 따르면 트리케라톱스 牙城體 (A, 트리케라톱스 프로르수스 완모式標本 YPM 1822)의 프릴이 길어지고 구멍이 생겨 B, 吐露社友루스 라투스 標本 ANSP 15192 가 되었을 것이다
 마지막 成長段階에서는 YPM 1831 標本 (A) 에서처럼 납작하고 巨大한 프릴을 볼 수 있었을 것이다. 막 成體가 된 ANSP 15192 (B) 와의 크기 比較
마지막 成長段階에서는 YPM 1831 標本 (A) 에서처럼 납작하고 巨大한 프릴을 볼 수 있었을 것이다. 막 成體가 된 ANSP 15192 (B) 와의 크기 比較
2010年에 스카넬라와 스카넬라의 指導敎授인
몬태나 州立 大學
의
잭 호너
는 헬크릭 層에서 發見된 38 個의 頭蓋骨 標本을 가지고 成長 패턴을 硏究하여 論文을 發表했다. 38 個의 標本 中 29 個는 트리케라톱스였고 9 個는 吐露社友루스였다. 스카넬라와 好너는 吐露사우루스가 트리케라톱스의 聖體 形態를 나타낸다고 結論내렸다.
[6]
好너는 角龍類 頭蓋骨의 프릴은 化生骨 (metaplastic bone) 로 이루어져 있다는 것을 强調했다. 化生골의 特徵은 時間이 지나면서 길어지거나 짧아질 수 있으며 늘어나거나 旣存의 뼈組織을 再吸收해 새로운 形態를 만들어낼 수 있다는 點이다. 트리케라톱스로 分類된 頭蓋骨들에서도 相當한 模樣 變化가 보인다고 好너는 이야기한다. "어릴 때는 뿔이 뒤로 굽어 있다가 成體가 되면 앞으로 휘어집니다." 大略 50% 程度의 牙城體 트리케라톱스 頭蓋骨은 吐露社友루스라면 구멍이 있었을 位置의 프릴 部位에 顆粒上 뼈로 둘러싸인 얇은 部位를 두 個 가지고 있다. 이런 形態는 트리케라톱스 個體가 成熟하면서 프릴이 길어져 늘어나는 무게를 相殺시키기 위한 것으로 보인다. 好너는 이것을 包含해 알려진 恐龍 종들이 實際로는 다른 種의 成長段階인 境遇가 많다는 大膽한 主張을 했다.
[27]
[28]
트리케라톱스의 나이 든 個體는 프릴이 相當한 比率로 길어지면서 납작해지고 뒤쪽 가장자리에서는 넓어졌을 것이다. 그와 同時에 마루뼈의 "구멍" 이 생겨나 典型的인 카스모社友루스아과의 프릴 形態를 만들었을 것이다.
[6]
스카넬라와 好너는 이 假說로 모든 資料를 깔끔하게 說明할 수 없다는 點은 認定했다. 이것을 說明하기 위해
補助假說
을 發展시켰다. 한 가지 問題는 吐露사우루스가 트리케라톱스의 正常的인 最後 成長段階, "吐露型"으로 불리는 形態라면 吐露社友루스 化石이 相當히 흔해야 하는데 그렇지 않다는 點이다. 스카넬라와 好너는 아성체의 높은 死亡率과 나이든 個體가 高地帶에서 살아
浸蝕
으로
化石化
가 드물게 일어났을 수 있다는 可能性 等으로 이것을 說明했다. 두番째 問題는 吐露社友루스 標本의 크기 分布를 보면 眞짜 吐露社友루스 아성체가 存在한다고 볼 수 있다는 點이다. 스카넬라와 好너는 뼈의 構造를 보면 該當 標本은 成長이 끝난 狀態이며 이런 크기의 差異는 個體 間의 變異에 依한 것으로 보인다고 主張했다. 세番째 問題는 마루뼈의 구멍이 恒常 完全한 形態로 存在해서 구멍이 있는 個體와 없는 個體 사이의 中間이 없는 것 같다는 點이다. 이에 對한 反論으로 스카넬라와 好너는 論爭거리인
네度케라톱스
의 완모式標本 USNM 2412 가 바로 그 中間 形態의 例라고 指摘한다. 이 속의 特徵들에 問題가 있는 것처럼 보이는 理由는 이것이 "吐露型"으로 轉換되는 첫番째 段階이기 때문이라는 것이다. 마지막 問題는 뒷머리뼈에 있는 프릴 가장자리 突起들의 數에 對한 것이다. 트리케라톱스는 普通 中心線에 있는 骨片을 包含해 마루뼈뒷突起가 다섯 個 있고, 吐露社友루스는 열 個 或은 열두 個 있으며 中心線의 마루뼈뒷突起는 存在하지 않는다. 프릴의 側面에 있는 비늘뼈뒷突起의 數도 달라서 트리케라톱스는 다섯 個, 吐露社友루스는 여섯 個나 일곱 個다. 이것은 뒷머리의 突起들 數字는 個體가 成熟함에 따라 늘어난다는 家庭을 하면 說明이 可能하다. 또 MOR 2923 은 여섯 個의 마루뼈뒷突起가 있는데 中心線에는 骨片이 없는 것을 보면 骨片의 位置나 數字는 트리케라톱스 안에서도 變異가 있다는 點이 指摘되었다.
[6]
스카넬라와 호너의 結論에 모두가 同意하는 것은 아니다. 여러 專門家들은 "吐露型" 假說이 맞을 可能性이 있다는 것은 認定하면서도 實際로 그랬을 可能性은 낮다고 主張했다. 이 假說에 對한 直接的인 反論은 2011年 앤드류 派키와 2012年 니콜라스 롱리치의 論文에서 提示되었다. 派키는 2011年에 問題가 되던
네度케라톱스 해處理
를 다시 記述하면서 이것이 스카넬라와 호너가 主張했던 것처럼 트리케라톱스가 아니라 獨立된 속으로 나이가 들어가 병든 個體라고 結論내렸다. 派키는 네度케라톱스의 프릴에 있는 不規則한 形態의 구멍은 뼈가 얇아지면서 뚫린 구멍이 아니며 오히려 구멍 周邊이 두꺼워져 있다는 點을 指摘했다. 派키는 또 트리케라톱스가 吐露社友루스로 成長한다는 假說과 맞아들어가지 않는 事實들이 여러 가지 있다고 結論내렸다. 全般的으로 케라톱스과에서 머리뼈 뒤쪽의 突起들은 프릴이 커지면서 늘어나지 않는다. 비늘뼈뒷突起의 數에 種種 變異가 있긴 하지만 어린 個體가 가장 많은 數의 突起를 가지고 있는 境遇도 있는 等 크기와는 相關이 있어 個體成長過程에서 變한다기보다는 個體 別로 있는 變異인 것으로 보인다. 이와 마찬가지로 全般的으로 각룡류에서 프릴의 구멍 形成은 나이와 相關이 없어서 가장 어린 個體라고 하더라도 마루뼈에 "구멍"을 가지고 있다. 트리케라톱스 프릴에 뼈가 얇은 部分, 卽 구멍이 생길 部分이라고 하는 部分은 派키의 說明에 따르면 筋肉이 附着되는 자리라고 한다. 또 구멍과 과립형 뼈 構造와는 一貫된 關係가 없다. 多數의 트리케라톱스 標本들이 프릴에 깊이 파인 血管 자국이 있는 것을 보면 나이가 많은 것이다. 이런 個體들의 프릴을 構成하는 뼈가 다시 젊어지고 과립형 뼈가 되어 구멍이 생기게 된다는 것은 派키가 보기에는 그럴법하지 않은 일이라는 것이다. 마지막으로 派키는 YPM 1831 標本은 그 엄청난 크기에도 不拘하고 封合되지 않은 뼈들이 있고 뼈의 表面組織이 매끈한 것을 보면 成熟이 끝나지 않은 것으로 보이며 따라서 眞正한 吐露社友루스의 어린 個體로 보인다는 點을 指摘했다.
[9]
 ANSP 15192 標本은 롱리치에 따르면 이제 막 成體가 된 암컷이라고 한다
ANSP 15192 標本은 롱리치에 따르면 이제 막 成體가 된 암컷이라고 한다
같은 해에 스카넬라와 好너는 派키의 批評에 對해 一部 答을 했다. 이들은 USNM 2412는 病理的인 狀態를 보면 中間形態로서는 그리 적합하지 않다는 點을 認定했지만 부풀어오른 部分을 除外하면 프릴의 구멍은 顆粒上으로 얇아진 뼈로 둘러싸여 있다는 點을 强調했다. 모든 證據를 考慮해 볼 때 스카넬라와 好너는 네度케라톱스가 獨立된 속이라기보다 病에 걸린 트리케라톱스 個體일 可能性이 높다고 主張했다. 또 이들은 트리케라톱스 標本들 中 派키가 不可能한 組合이라고 말한 血管 자국이 있는 顆粒上의 어린 뼈가 있다는 點을 指摘했다. 스카넬라와 好너는 또 트리케라톱스 프릴의 얇은 部分에 筋肉이 附着되었다는 主張을 그 部分이 筋肉附着面에서 一般的으로 보이는 特徵인 주름진 表面이 아니라는 理由로 棄却했다. 프릴 周圍 突起들의 數에 對해서는 두 가지 追加 說明을 내놓았다. 個體들이 나이 먹으면서 骨片이 닳아 마치 하나가 두 個인 것처럼 보일 수 있다는 것이다. 이 境遇 다섯 個 或은 여섯 個의 트리케라톱스 마루뼈뒷突起는 吐露社友루스 標本에서 볼 수 있는 것처럼 熱에서 열두 個까지 늘어날 수 있다. 다른 說明으로는 吐露社友루스 라투스 標本들은 더 오래된 地層에서 發見되기 때문에 아나제네시스(anagenesis) 過程, 卽 트리케라톱스 進化의 初期 段階에 있었을지 모른다는 것이다. 가장 오래된 標本인 MOR 1122는 열두 個의 마루뼈뒷突起를 가지고 있고 가장 最近의 標本인 MOR 981 銀 열 個를 가지고 있어 마루뼈뒷突起의 數가 줄어드는 一連의 進化過程을 나타내는 것이라고 볼 수도 있다.
[29]
2012年에 롱리치는
反證可能性
의 原理를 適用하여 이 問題를 調査했다. 有效한 科學的 假說은
豫測
을 하며 이 豫測은
檢證
될 수 있다. 롱리치는 "吐露型" 假說에서 세 個의 豫測을 이끌어낼 수 있다고 主張했다. 첫째로, 萬一 吐露사우루스가 트리케라톱스라면 이 둘의 化石은 같은 場所에서 發見되어야 한다. 實際로 이들의 地理的 分布範圍는 完全히 겹치지는 않는다. 北쪽으로 멀리 가면 吐露社友루스 化石은 發見되지 않는데 비해 南쪽으로 내려가면 吐露社友루스 右打헨시스만이 알려져 있다. 하지만 이것은 相對的으로 드문 吐露사우루스가 덜 發見되기 때문에 나타나는 錯視現象日 수 있다. 롱리치는 따라서 첫番째 豫測은 假說을 支持한다고 보았다. 두番째 豫測은 吐露社友루스 標本은 모두 聖體여야 하며 트리케라톱스 標本은 많이 늙은 個體가 없어야 한다. 롱리치에 따르면 이 問題는 完全히 解決되지 않았다고 한다. 2011年에 호너가 骨組織學 硏究를 發表하면서 調査에 利用된 트리케라톱스 標本이 아성체의 뼈 構造를 가지고 있다는 것을 보였지만,
[30]
標本의 크기가 너무 작아서 모든 트리케라톱스 化石이 다 그렇다고 一般化시키기는 無理가 있다. 이 豫測을 더 잘 檢證하기 위해 롱리치는 24 가지 頭蓋骨의 特徵 目錄을 提案했다. 이 目錄을 가지고 標本을 調査하면 頭蓋骨의 여러 뼈들이 하나로 합쳐지는 成熟度를 測定할 수 있다. 이 基準에 따라 36 個의 標本을 調査했다. 뼈가 합쳐지는 過程은 普通 특정한 順序로 進行되어 나이에 對한 追加情報를 얻을 수 있다. 實際로 이 基準에 따르면 大部分의 吐露社友루스 標本은 매우 늙은 個體였다. 하지만 두 가지 例外가 있었다. 작은 個體인 ANSP 15192는 聖體긴 했지만 주둥이의 뼈가 합쳐지지 않은 것으로 보아 相對的으로 젊은 個體였다. 가장 어린 標本은 YPM 1831 이었는데 주둥이, 광대뼈의 뿔 과 後頭과 等이 합쳐지지 않은 狀態였다. 게다가 프릴의 骨片들이 없는 것으로 보아 이들도 합쳐지지 않은 것으로 보이며 프릴의 가장자리는 겉보기에 如前히 成長 中인, 어린 뼈인 것으로 보인다. 그 反面에 롱리치는 열 個의 트리케라톱스 頭蓋骨을 調査해 가장 나이가 많은 吐露社友루스 標本과 같은 程度의 成熟度를 보인다는 것을 알아냈다. 롱리치는 두番째 豫測의 檢證結果는 "吐露型" 假說을 反證한다고 結論내렸다. 세番째 豫測은 吐露社友루스와 트리케라톱스 사이의 中間形態가 있어야 한다는 것이다. 롱리치는 트리케라톱스 프릴의 얇은 部分이 마루뼈 "구멍" 이 形成되는 過程이라는 主張이 가장 强力한 中間形態의 證據라고 보고 이 主張을 살펴보았다. 하지만 롱리치는 이 救助의 位置가 다르다는 點을 指摘했다. 트리케라톱스의 움푹 들어간 部分은 一部가 비늘뼈에 걸쳐 있으나 吐露社友루스의 구멍은 마루뼈 가운데 있다. 게다가 움푹 들어간 部分의 가장자리에 있는 뼈는 더 두꺼워져 있는데 비해 吐露社友루스의 구멍은 얇은 뼈로 둘러싸여 있었다. 롱리치는 "吐露型" 假說이 세番째 豫測에서도 反證된다고 結論내렸다. 세 個의 豫測 가운데 두 個의 豫測이 假說을 反證하므로 "吐露型" 假說은 棄却되어야 한다.
[4]
롱리치는 또 "吐露型" 假說에 對한 追加 反駁을 提示했다. 마루뼈뒷突起의 數와 關聯해서 中間形態가 存在하지 않는다. 또 이 突起들의 프릴의 가장자리를 꽉 채우고 있는데 어떻게 그 數字가 늘어날 수 있는지도 알기 힘들다. 化生骨 狀態로 成長한다면 骨片들의 크기가 프릴과 함께 커질 것이기 때문이다. 骨片이 닳아서 두 個로 나뉜다는 主張은 비늘뼈뒷突起에 對해서만 適用되고 마루뼈뒷突起와는 相關이 없다. 吐露社友루스의 비늘뼈는 안쪽이 두껍고 바깥쪽 表面이 오목한 反面 트리케라톱스의 비늘뼈는 안쪽이 오목하고 위쪽은 扁平하다. 中間形態는 알려지지 않았다. 吐露社友루스의 비늘뼈는 또 絶對的인 크기와 相關없이 더 길쭉하다. 롱리치는 吐露社友루스와 트리케라톱스 標本들이 스카넬라와 호너의 主張처럼 하나의 成長段階라고 본다면 吐露社友루스 標本인 ANSP 15192 와 YPM1831 은
相對成長
으로 說明할 수 있는 것보다 더 긴 形態로 回歸線에서 벗어난 아웃라이어가 된다는 點을 指摘했다. 롱리치는 호너가 트리케라톱스의 牙城體만을 볼 수 있었다는 것이 吐露型 假說을 支持하는 것으로 보일 수 있다고 認定했지만 트리케라톱스가 近緣種들보다 어린 뼈의 構造를 나이 들어서까지 維持하고 있는 것으로 볼 수도 있다고 主張했다. 또 한 篇, 뼈의 再모델링으로 成熟度를 評價하는 것은 信賴할 만한 方法이 아니다. 實驗을 통한 硏究를 보면 다양한 種類의 뼈에 加해진 疫學的인 壓力 條件의 差異에 따라 이런 再모델링 程度가 크게 變할 수 있으며 오래된 뼈 組織인 것처럼 보이는 것이 可能하다. 롱리치는 스카넬라와 호너가 自身의 두番째 檢證 內容에 對해 個體變異 때문에 생긴 結果라고 反駁할 것이라고 豫測했다. 롱리치에 따르면 個體變異라는 要因의 重要性은 制限的이라고 한다. ANSP 15192 와 YPM 1831 은 그보다
成績 이형성
으로 더 잘 說明될 수 있으며 ANSP 15192는 젊은 聖體 암컷, 그리고 YPM 1831 은 牙城體 수컷으로 볼 수 있다고 主張했다.
[4]
 頭蓋骨(A)와 비늘뼈(B)에 對한 主成分分析(PCA)와 線型回歸分析 結果. 吐露社友루스는 크기와 無關하게 트리케라톱스 호리두스 및 트리케라톱스 프로르수스와 따로 떨어진 形態空間을 차지하고 있다
頭蓋骨(A)와 비늘뼈(B)에 對한 主成分分析(PCA)와 線型回歸分析 結果. 吐露社友루스는 크기와 無關하게 트리케라톱스 호리두스 및 트리케라톱스 프로르수스와 따로 떨어진 形態空間을 차지하고 있다
2013年에 派키와 레오나르도 마이오리노는 吐露社友루스, 트리케라톱스 호리두스, 트리케라톱스 프로르수스, 그리고 네度케라톱스의 頭蓋骨과 成熟度에 따른 形態의 變異를 보여주는
形態空間
에 對한 統計的 分析을 遂行한
形態測定學
硏究를 出版했다. 이들은 吐露社友루스 라투스 頭蓋骨은 成熟過程 내내
T. horridus
및
T. prorsus
와는 다른 形態를 維持하는 反面
T. horridus
와
T. prorsus
는 比率 面에서 形態가 겹친다고 結論내렸다. 프릴의 模樣을 除外한다고 하더라도 이 結論은 成立한다. 네度케라톱스는 크기를 除外하면 吐露社友루스와 트리케라톱스 호리두스의 中間形態라고 보기 힘들다. 派키와 마이오리노는 吐露社友루스 標本의 數가 적기 때문에 結果의 信賴性이 減少한다는 點은 認定했지만 아나제네시스의 可能性을 勘案한다 하더라도 吐露社友루스와 트리케라톱스는 別個의 分類群이라는 結論을 내렸다.
[10]
吐露社友루스 라투스 標本이 트리케라톱스 成熟 過程의 "吐露型" 段階를 보여준다는 假說은 두番째 吐露社友루스 종인 吐露社友루스 우타헨시스 亦是 吐露兄의 하나가 아닌가 하는 疑問을 提起했다. 이 問題는 大槪의 標本이 따로 떨어져 發見된 뼈라서 保存狀態가 좋은 標本이 없다는 點 때문에 더 複雜해진다.
T. utahensis
가 吐露社友루스로 分類되는 주된 理由는 길쭉한 비늘뼈, 卽 긴 프릴 때문이다. 마루뼈뒷突起의 數와 크기, 位置, 게다가 마루뼈의 "구멍" 이 있는지도 알려지지 않았다. 어린 吐露사우루스가 빅벤드 國立公園의 자벨리나 層의 뼈層에서 發掘되었다는 主張이 있다. 聖體가 發見된 位置와 가까운 곳이고 吐露社友루스 마루뼈의 特徵을 보아 이 標本은 吐露社友루스 cf. 右打헨시스로 分類하고 있다.
[19]
스카넬라와 好너는 向後에 標本들이 追加로 發見되어야만 問題가 解決된다고 結論내리고 있다. 이들은 吐露社友루스의 棲息範圍를 트리케라톱스의 範圍보다 더 南쪽으로 넓히게 되는데, 이것이 카스모社友루스亞科에 屬하는 別途의 속이거나 트리케라톱스의 세番째 終日 可能性도 있다고 보고 있다. 派키는 2013年의 形態測定學 硏究는 이런 點에서 明確하지 않은 結果를 보여주고 있는데,
T. utahensis
의 形態空間이 트리케라톱스와 吐露社友루스 라투스의 中間에 位置하고 있으며 兩쪽 모두로부터 뚜렷이 分離되어 있지 않다.
[10]
古生物學
[
編輯
]
吐露社友루스와 트리케라톱스를 包含하여 모든 각룡류는
草食動物
이었다. 턱에는 여러 줄의 이빨이 있고 턱이 닫히면서 위아래 이빨이 빗겨지나가게 되어 있어 질긴 植物을 잘라먹을 수 있었다. 아마 大槪의 땅을 덮고 있는 羊齒類들을 主로 먹었을 것이다. 트리케라톱스보다 吐露社友루스의 주둥이가 더 길고 좁았던 것으로 보아 食性에는 差異가 있었던 것으로 보인다.
[
出處 必要
]
各州
[
編輯
]
- ↑
Paul, G.S.; Christiansen, P. (2000).
“Forelimb posture in neoceratopsian dinosaurs: implications for gait and locomotion”
(PDF)
. 《Paleobiology》
26
(3): 450?465.
doi
:
10.1666/0094-8373(2000)026[0450:FPINDI]2.0.CO;2
(年 以後로 接續 不可 2022-08-01).
- ↑
Paul, G. S.
(2010).
《The Princeton Field Guide to Dinosaurs》
. Princeton University Press.
265?267
쪽.
ISBN
978-0-691-13720-9
.
- ↑
Holtz, Thomas R. Jr. (2012).
《Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages》
(PDF)
.
Winter 2011 Appendix
- ↑
가
나
다
라
마
Longrich, N. R and Field, D. J. (2012).
“
Torosaurus
is not
Triceratops
: Ontogeny in chasmosaurine ceratopsids as a case study in dinosaur taxonomy”
.
PLoS ONE
7
(2): e32623.
Bibcode
:
2012PLoSO...7E2623L
.
doi
:
10.1371/journal.pone.0032623
.
PMC
3290593
.
PMID
22393425
.
- ↑
“Morph-osaurs: How shape-shifting dinosaurs deceived us - life - 28 July 2010”
. New Scientist.
doi
:
10.1080/02724634.2010.483632
. 2010年 8月 3日에 確認함
.
- ↑
가
나
다
라
마
Scannella, J. and Horner, J.R. (2010). "
Torosaurus
Marsh, 1891, is
Triceratops
Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny ."
Journal of Vertebrate Paleontology
,
30
(4): 1157 - 1168.
doi
10.1080/02724634.2010.483632
- ↑
Switek, Brian.
“New Study Says Torosaurus=Triceratops”
.
Dinosaur Tracking
. Smithsonian.com. 2013年 11月 13日에
原本 文書
에서 保存된 文書
. 2011年 3月 2日에 確認함
.
- ↑
Horner, Jack. TEDX Talks: "
Shape-shifting Dinosaurs
Archived
2014年 2月 16日 -
웨이백 머신
". Nov 2011. Accessed 20 Nov 2012.
- ↑
가
나
Farke, A. A. (2011) "
Anatomy and taxonomic status of the chasmosaurine ceratopsid
Nedoceratops hatcheri
from the Upper Cretaceous Lance Formation of Wyoming, U.S.A..
"
PLoS ONE
6
(1): e16196.
doi
10.1371/journal.pone.0016196
- ↑
가
나
다
Maiorino L., Farke A.A., Kotsakis T., Piras P.,2013, "Is
Torosaurus
Triceratops
? Geometric Morphometric Evidence of Late Maastrichtian Ceratopsid Dinosaurs",
PLoS ONE
8
(11): e81608. doi:10.1371/journal.pone.0081608
- ↑
가
나
O.C. Marsh, 1891, "Notice of new vertebrate fossils",
The American Journal of Science, series 3
42
: 265-269
- ↑
Dodson, P.
The Horned Dinosaurs
. Princeton Univ. Press (Princeton), 1996.
- ↑
Colbert, E.H. and J.D. Bump, 1947, "A skull of
Torosaurus
from South Dakota and a revision of the genus:,
Proceedings of the Academy of Natural Sciences of Philadelphia
,
99
: 93?106
- ↑
Farke, A., 2002, "A review of
Torosaurus
(Dinosauria: Ceratopsidae) specimens from Texas and New Mexico",
Journal of Vertebrate Paleontology
,
22
: 52A
- ↑
Lucas, S.G., Mack, G.H., Estep, G.W., 1998, "The Ceratopsian dinosaur
Torosaurus
from the Upper Cretaceous McRae Formation, Sierra County, New Mexico",
New Mexico Geological Society Guidebook, 49th Field Conference, Las Cruces County II
- ↑
C.W. Gilmore, 1946, "Reptilian fauna of the North Horn Formation of central Utah",
United States Department of the Interior Geological Survey Professional Paper
210
-C: 29-53
- ↑
D.A. Lawson, 1976, "
Tyrannosaurus
and
Torosaurus
, Maestrichtian dinosaurs from Trans-Pecos, Texas",
Journal of Paleontology
50
(1): 158-164
- ↑
Sullivan, R. M., A. C. Boere, and S. G. Lucas. 2005. Redescription of the ceratopsid dinosaur
Torosaurus utahensis
(Gilmore, 1946) and a revision of the genus.
Journal of Paleontology
79:564-582.
- ↑
가
나
다
Hunt, Rebecca K. and Thomas M. Lehman. 2008. Attributes of the ceratopsian dinosaur
Torosaurus
, and new material from the Javelina Formation (Maastrichtian) of Texas. Journal of Paleontology 82(6): 1127-1138.
- ↑
Hatcher, J.B., Marsh O.C. and Lull, R.S., 1907,
The Ceratopsia
, Monographs of the United States Geological Survey
49
: 1-198
- ↑
Lull, R.S., 1933,
A revision of the Ceratopsia or horned dinosaurs
, Memoirs of the Peabody Museum of Natural History
3
(3): 1-175
- ↑
Lehman, T.M., 1998, "A gigantic skull and skeleton of the horned dinosaur
Pentaceratops sternbergi
from New Mexico: Journal of Paleontology,
72
(5): 894-906
- ↑
Nicholas R. Longrich, 2011, "
Titanoceratops ouranous
, a giant horned dinosaur from the Late Campanian of New Mexico",
Cretaceous Research
,
32
: 264-276
- ↑
가
나
다
라
마
Farke, A. A. "Cranial osteology and phylogenetic relationships of the chasmosaurine ceratopsid
Torosaurus latus
", pp. 235-257. In K. Carpenter (ed.).
Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs
. Indiana Univ. Press (Bloomington), 2006.
- ↑
가
나
Scott D. Sampson, Mark A. Loewen, Andrew A. Farke, Eric M. Roberts, Catherine A. Forster, Joshua A. Smith, and Alan A. Titus (2010).
“New Horned Dinosaurs from Utah Provide Evidence for Intracontinental Dinosaur Endimism”
.
PLoS ONE
. 5
5
(9): e12292.
doi
:
10.1371/journal.pone.0012292
.
PMC
2929175
.
PMID
20877459
.
- ↑
Scannella J., 2009, "And then there was one: synonymy consequences of
Triceratops
cranial ontogeny",
Journal of Vertebrate Paleontology
29
: 177A
- ↑
“New Analyses Of Dinosaur Growth May Wipe Out One-third Of Species”
. Sciencedaily.com. 2009年 10月 31日
. 2010年 8月 3日에 確認함
.
- ↑
doi
10.1371/journal.pone.0007626
現在 이 引用은 內容이 不完全합니다.
英語版의 文書에서 複寫
하여 完成할 수 있습니다.
- ↑
Scannella, J.B. & Horner, J.R., 2011, "‘Nedoceratops’: An Example of a Transitional Morphology",
PLoS ONE
6
(12): DOI:10.1371/journal.pone.0028705
- ↑
Horner, J.R., Lamm, E-T., 2011, "Ontogeny of the parietal frill of
Triceratops
: a preliminary histological analysis",
Comptes Rendus de l’Academie des Sciences Paris serie D
10
: 439?452
- Dodson, P. (1996).
The Horned Dinosaurs
. Princeton University Press, Princeton, New Jersey, pp. xiv-346
外部 링크
[
編輯
]