偏母
(flagellum, flagella)는 運動性을 提供하기 위해 特定
植物
과 動物의 精子 細胞와 다양한
微生物
이 가진 突出된 털이 있는 附屬 機關이다.
[1]
[2]
[3]
[4]
鞭毛가 있는 많은
原生生物
을
鞭毛蟲類
라고 한다.
微生物은 하나에서 많은 鞭毛를 가질 수 있다. 例를 들어
그람 음성균
人
헬리코박터 파일로리
(
Helicobacter pylori
)는 多衆 鞭毛를 利用하여 粘液 內壁에 닿아 位
上皮
에 到達하여
胃潰瘍
을 誘發할 수 있다.
[5]
一部 박테리아에서 鞭毛는 細胞 外部의 濕氣에 敏感한 感覺
小器官
으로도 기능할 수 있다.
[6]
細菌
,
古筠
,
眞核生物
의
歲 驛
에 걸쳐 鞭毛는 救助, 蛋白質 構成 및 推進 메커니즘이 다르지만 運動性 提供이라는 同一한 機能을 共有한다.
라틴語
flagellum(flagellum)는
채찍
을 의미하여 채찍 같이 헤엄치는 動作을 의미한다. 高菌의 鞭毛는 細菌 鞭毛와의 差異點을 記錄하기 위해 archaellum이라고 한다.
[7]
[8]
眞核生物의 偏母와 纖毛는 構造가 同一하지만 길이와 機能이 다르다.
[9]
原核生物
의
仙母
는 더 작고 얇은 附屬機關으로 機能이 다르다.
類型
[
編輯
]
 原核生物
(細菌과 古筠) 鞭毛는 回轉 運動으로 움직이는 反面, 眞核生物 鞭毛는 굽힘 運動으로 움직인다. 原核生物 鞭毛는 回戰 모터를 使用하고 眞核生物 鞭毛는 複雜한 슬라이딩 필라멘트 시스템을 使用한다. 眞核生物의 鞭毛는 ATP에 依해 움직이는 反面, 原核生物의 鞭毛는 ATP에 依해 움직이거나(고균) 陽性子에 依해움직일(세균) 수 있다.
[10]
原核生物
(細菌과 古筠) 鞭毛는 回轉 運動으로 움직이는 反面, 眞核生物 鞭毛는 굽힘 運動으로 움직인다. 原核生物 鞭毛는 回戰 모터를 使用하고 眞核生物 鞭毛는 複雜한 슬라이딩 필라멘트 시스템을 使用한다. 眞核生物의 鞭毛는 ATP에 依해 움직이는 反面, 原核生物의 鞭毛는 ATP에 依해 움직이거나(고균) 陽性子에 依해움직일(세균) 수 있다.
[10]
鞭毛의 세 가지 類型은 細菌, 古筠, 眞核生物이다.
眞核生物의 鞭毛에는 굽힘 메커니즘으로 움직이는
디네인
과
微細小管
이 있다. 박테리아와 古細菌은 偏母에 디네인이나 微細小管이 없으며 回戰 메커니즘을 使用하여 움직인다.
[11]
이 세 가지 類型의 다른 差異點은 다음과 같다.
- 細菌의 鞭毛는 螺旋形 필라멘트이며, 各各은 時計 方向 또는 反時計 方向으로 回轉할 수 있는 回轉 모터가 바닥에 있다.
[12]
[13]
[14]
回戰 모터를 통해 여러 種類의 박테리아 運動性 中 두 가지가 나타난다.
[15]
[16]
- 高菌의 鞭毛(archaella)는 回戰 모터를 가지고 있다는 點에서 表面的으로 細菌 偏母와 類似하지만 많은 細部的인 特徵은 다르며 非
同種
으로 看做된다.
[17]
[18]
[19]
- 眞核生物 鞭毛(動物, 植物, 原生生物 細胞의 鞭毛)는 앞뒤로 휘몰아치는 複雜한 細胞 突出部이다. 眞核生物의 偏母와 運動性 纖毛는 構造가 同一하지만 길이, 波形, 機能이 다르다.
- 1次 纖毛는 運動性이 없으며 偏母와 運動性 纖毛 모두에서 發見되는 9+2 軸索과는 달리 構造的으로 다른 9+0 軸索을 가지고 있다.
細菌
[
編輯
]
構造 및 構成
[
編輯
]
細菌性 鞭毛는 鞭毛의
蛋白質
小段位로 構成된다.
[11]
그 模樣은 20
나노미터
두께의 속이 빈 튜브이다.
螺旋形
이며 外膜 바로 바깥쪽에 날카로운 屈曲이 있다. 이 "후크"를 使用하면 나선의 軸이 細胞에서 直接 멀어지도록 할 수 있다. 샤프트는 후크와 基底體 사이를 달리며 베어링 役割을 하는 細胞膜의 蛋白質 고리를 通過한다.
그람 陽性
有機體는
펩티도글리칸
層에 하나,
原形質膜
에 하나, 이러한 基礎 몸體 고리 中 2個를 가지고 있다.
그람 陰性
有機體는 4個의 이러한 고리를 가지고 있다. L 고리는
吏逋폴리사카라이드
와 結合하고, P 고리는
펩티도글리칸
層과 結合하고, M 고리는
原形質膜
에 內臟되고, S 고리는 原形質膜에 直接 附着된다. 필라멘트는 캡핑 蛋白質로 끝난다.
[20]
[21]
偏母 필라멘트는 모터에 依해 回傳될 때 후크를 通해 박테리아를 推進하는 긴 螺旋形 螺絲이다. 그람 陰性
Escherichia coli, Salmonella typhimurium, Caulobacter crescentus
,
Vibrio alginolyticus
를 包含하여 硏究된 大部分의 박테리아에서 필라멘트는 필라멘트 軸에 大略 平行한 11個의 原形 필라멘트로 構成된다. 各 原形 필라멘트는 一連의 2引用 自轉車(탠덤, tandem) 蛋白質 사슬이다. 그러나
Campylobacter jejuni
에는 7個의 原形纖維가 있다.
[22]
基底體는 原形質膜을 통해 뻗어 있는 中央에 있는 속이 빈 막대 模樣의 "플러그"와 같은 一部 類型의
分泌 구멍
과 共通되는 몇 가지 特性을 가지고 있다. 細菌 偏母와 細菌 分泌 시스템 構造 및 蛋白質 間의 類似性은 細菌 鞭毛가 3型 分泌 시스템에서 進化했다는 理論을 뒷받침하는과학적 證據를 提供한다.
모터
[
編輯
]
박테리아 鞭毛는 內部 細胞膜의 鞭毛 固定點에 位置한 蛋白質로 構成된 回戰 엔진(Mot complex)에 依해 구동된다. 엔진은
陽性子 驅動力
, 卽 細胞의 新進 臺詞에 依해 設定된 濃度 勾配로 因해 박테리아 細胞膜을 가로 지르는 陽性子(水素 이온)의 흐름에 依해 구동된다(
Vibrio
鐘에는 側面 및 極盛의 두 種類의 鞭毛가 있다. 그리고 一部는
陽性子 펌프
가 아닌 나트륨
이온 펌프
에 依해 구동된다
[23]
). 로터는 膜을 가로질러 陽性子를 運搬하고 그過程에서 回轉한다.
로터 單獨으로 6,000-17,000rpm으로 作動할 수 있지만 偏母 필라멘트가 附着된 境遇 一般的으로 200-1000
rpm
에 到達한다. 回轉 方向은 로터에서 蛋白質 FliG의 位置가 若干 變更되어 거의 瞬間的으로 鞭毛 모터 스위치에 依해 變更될 수 있다.
[24]
鞭毛는 에너지 效率이 높고 에너지를 거의 使用하지 않는다.
[25]
토크 生成에 對한 正確한 메커니즘은 아직 제대로 理解되지 않았다.
[26]
偏母 모터에는 온-오프 스위치가 없기 때문에 蛋白質 epsE는 모터를 로터에서 分離하는 機械的 클러치로 使用되어 鞭毛를 멈추고 박테리아가 한 곳에 남아 있게 한다.
[27]
鞭毛의 圓筒形 模樣은 微細한 有機體의 移動에 適合하다. 이 有機體는 周邊 물의 粘度가 質量이나 慣性보다 훨씬 더 重要한 낮은
레이놀즈 수
에서 作動한다.
[28]
鞭毛의 回轉 速度는 陽性子 動力의 强度에 따라 달라지므로 特定 形態의 速度 制御가 可能하고 一部 類型의 박테리아는 크기에 比例하여 놀라운 速度를 얻을 수 있다. 一部는 秒當 大略 60個의 셀 길이를 達成한다. 이러한 速度로 박테리아가 1km를 덮는 데 約 245日이 걸린다. 느린 것처럼 보일 수 있지만 規模의 槪念이 導入되면 觀點이 바뀐다. 巨視的인 生命體와 比較할 때 秒當 몸 길이의 數로 表現하면 實際로 매우 빠르다. 例를 들어, 치타는 秒當 約 25個의 몸 길이 程度만을 달릴 수 있다.
[29]
鞭毛의 使用을 통해 박테리아는
偏向된 無作爲 步行
을 통해 油印物 쪽으로 빠르게 移動하고 忌避劑에서 멀리 移動할 수 있으며 鞭毛를
時計 反對 方向
및
時計 方向
으로 各各 回轉하여 '달리기' 및 '뒤집기'를 遂行한다. 두 回轉 方向은 同一하지 않으며(편모 運動과 關聯하여) 分子 스위치에 依해 選擇된다.
[30]
]
組立
[
編輯
]
偏母 組立 中에 鞭毛의 構成 要素는 基底體의 中共 코어와 初期 필라멘트를 通過한다. 組立하는 동안 蛋白質 成分은 基底部보다는 鞭毛 末端에 追加된다.
[31]
試驗管 內에서 鞭毛 필라멘트는 精製된 鞭毛를 唯一한 蛋白質로 包含하는 溶液에서 自發的으로 組立된다.
[32]
鎭火
[
編輯
]
박테리아 鞭毛의 적어도 10個 蛋白質 成分은 많은 그람 陰性 박테리아에서 發見되는 3型 分泌 시스템(T3SS)과 상동 蛋白質을 共有하므로
[33]
하나가 다른 것으로 鎭火했을 可能性이 있다. T3SS는 鞭毛 裝置와 類似한 數의 構成 要素(藥 25個의 蛋白質)를 가지고 있기 때문에 어떤 것이 먼저 進化했는지 決定하기 어렵다. 그러나 鞭毛 시스템은 多樣한 조절자와 샤페론을 包含하여 全般的으로 더 많은 蛋白質을 包含하는 것으로 보이므로 鞭毛가 T3SS에서 鎭火했다고 主張되어 왔다. 그러나 鞭毛가 먼저 進化했거나 두 構造가 竝列로 鎭火했을 수도 있다고 提案되었다.
[34]
初期 單細胞 有機體의 運動性(移動性)에 對한 要求는 進化에 依해 더 移動性이 높은 鞭毛가 먼저 選擇될 것임을 支持하지만,
[34]
鞭毛에서 進化하는 T3SS는 '還元的 進化'로 볼 수 있으며,
系統 發生
나무.
[35]
두 構造가 共通 祖上과 別途로 進化했다는 假說은 두 構造 사이의 蛋白質 類似性과 機能的 多樣性을 說明한다.
[36]
偏母와 知的 設計 論爭
[
編輯
]
一部 著者들은 鞭毛가 모든 蛋白質이 제자리에 있을 때만 제대로 기능할 수 있다고 假定하고 進化할 수 없다고 主張했다. 卽, 鞭毛 裝置는 "
還元 不可能하게 複雜
"하다.
[37]
그러나 많은 蛋白質이 削除되거나 突然變異될 수 있으며 鞭毛는 때때로 效率性이 떨어지더라도 如前히 作動한다.
[38]
또한, 種에 걸쳐 몇 가지 固有한 蛋白質이 많기 때문에 박테리아 鞭毛 造成의 多樣性은 豫想보다 높았다.
[39]
따라서 鞭毛 裝置는 進化論的 側面에서 分明히 매우 柔軟하며 蛋白質 成分을 完璧하게 잃거나 얻을 수 있다. 例를 들어, 大腸菌의 運動性을 증가시키는 多數의 突然變異가 發見되었다.
[40]
박테리아 鞭毛의 進化에 對한 追加 證據에는 痕跡 鞭毛의 存在, 鞭毛의 中間 形態 및 鞭毛 蛋白質 序列 間의 類似性 패턴이 包含되며, 여기에는 거의 모든 核心 鞭毛 蛋白質이 非鞭毛 蛋白質과의 相同性이 알려져 있다는 觀察이 包含된다.
[33]
또한, 單純 反復 서브유닛의 自家 組立, 後續 分期를 통한 遺傳子 複製, 다른 시스템의 要素 募集('分子 브리콜라주') 및 再組合을 包含하여 鞭毛 進化에서 重要한 役割을 하는 여러 프로세스가 確認되었다.
[41]
偏母 配列
[
編輯
]
박테리아의 種에 따라 鞭毛의 數와 配列이 다르다.
[42]
[43]
그리스어로
머리카락
을 의미하는
trichos
에서 따온
tricho
라는 用語를 使用하여 命名되었다.
[44]
- 콜레라菌
과 같은
Monotrichous bacteria
는 單一
極盛 鞭毛
를 가지고 있다.
[45]
- Amphitrichous bacteria
는 兩쪽 끝에 各各 하나의 鞭毛를 가지고 있다(예:
Alcaligenes faecalis
). 한 番에 하나의 片貌만 作動하므로 어느 鞭毛가 活性化되어 있는지 轉換하여 박테리아가 빠르게 進路를 바꿀 수 있다.
- Lophotrichous
bacteria
는
[46]
位螺旋菌
와 같이 박테리아 表面의 同一한 地點에 여러 個의 鞭毛가 있으며 박테리아를 한 方向으로 몰아가기 爲해 協力한다. 많은 境遇에, 여러 鞭毛의 基底部는 極盛 小器官이라고 하는 細胞膜의 特殊 領域으로 둘러싸여 있다.
- Peritrichous bacteria
는 모든 方向으로 突出된 鞭毛를 가지고 있다(예:
E. coli
).
Monotrichous bacteria의 鞭毛가 反時計 方向으로 回轉하면 코르크 內部에서 움직이는 코르크 마개처럼 鞭毛가 뒤에 오는 狀態로 細胞가 앞으로 밀려난다. 微視的 規模의 물은 一般
물
과 달리
粘性
이 매우 높다.
이에 反해 스피로헤타는 細胞의 反對 劇에서 發生하는 鞭毛 內幕(endoflagella)이라고 하는 鞭毛를 가지고 있으며, 外膜을 破壞하고 electron cryotomography microscope에 依해 보여지는 바와 같이 周邊 細胞質 空間 內에 位置한다.
[47]
[48]
[49]
細胞體에 對한 필라멘트의 回轉은 一般的으로 鞭毛가 있는 박테리아의 通過를 防止할 수 있을 만큼 充分히 粘性이 있는 物質을 통해서도 全體 박테리아가 코르크 마개 같은 움직임으로 앞으로 移動하도록 한다.
셀레老모나스
의 特定 大型 形態에서는 30個 以上의 個別 鞭毛가 細胞體 外部에 組織되어 있으며 서로에 對해 螺旋形으로 꼬여 "다낭"이라고 하는 두꺼운 救助(狂顯微鏡으로 쉽게 볼 수 있음)를 形成한다.
一部
Vibrio
spp. (特히
腸炎비브리오菌
[50]
),
에로모나스
와 같은 關聯
박테리아
, 두 個의 鞭毛 시스템이 共存하며 서로 다른 遺傳子 세트와에너지에 對한 서로 다른 이온 勾配를 使用한다. 極盛 鞭毛는 體液에서 構成的으로 發現되고 運動性을 提供하는 反面, 側面 鞭毛는 極 鞭毛가 回戰에 너무 많은 抵抗을 만날 때 發現된다.
[51]
[52]
[53]
[54]
[55]
[56]
이들은 表面이나 粘性 流體에서 무리지어 움직이는 運動性을 提供한다.
번들링
[
編輯
]
번들링(Bundling)은 鞭毛가 여러 個 있는 細胞에서 發生할 수 있는 이벤트로, 鞭毛를 함께 묶어 調整된 方式으로 回轉하게 한다.
鞭毛는 왼손 螺旋이며 로터에 依해 反時計 方向으로 回轉하면 함께 묶이고 回轉할 수 있다. 로터가 方向을 바꾸어 時計 方向으로 回轉하면 鞭毛가 번들에서 풀린다. 이로 인해 細胞가 前震 動作을 멈추고 代身 제자리에서 痙攣을 始作할 수 있다. 이를 "텀블링(tumbling)"이라고 한다. 텀블링은 細胞의 確率的 再配享을 招來하여 앞으로 헤엄치는 方向을 變更하게 한다.
어떤 刺戟이 번들링과 텀블링 사이의 스위치를 驅動하는지 알 수 없지만 모터는 다양한 信號에 매우 잘 適應한다.
走化性
("故意的인 움직임"을 說明하는 모델에서 鞭毛의 時計 方向 回轉은 細胞에 有利한 化合物(예: 飮食)에 依해 抑制된다. 유리한 方向으로 움직일 때, 그러한 化學 誘引 物質의 濃度가 增加하고 따라서 텀블이 持續的으로 抑制되어 앞으로 움직일 수 있다. 마찬가지로, 細胞의 運動 方向이 바람직하지 않을 때(例를 들어, 化學的 誘引 物質로부터 멀리 떨어져 있는 境遇), 텀블은 더 以上 抑制되지 않고 훨씬 더 자주 發生하여 細胞가 올바른 方向으로 再配享될 可能性이 있다.
그러나 모든 鞭毛가 時計 方向으로 回轉하더라도 幾何學的 및 流體 力學的 理由로 인해 번들을 形成하지 못하는 境遇가 많다.
[57]
[58]
眞核生物
[
編輯
]
 眞核生物 鞭毛. 1-axoneme, 2-細胞膜, 3-IFT(鞭毛內 輸送), 4-基底體, 5-鞭毛의 斷面, 6-基底體의 微細小管의 三重體
眞核生物 鞭毛. 1-axoneme, 2-細胞膜, 3-IFT(鞭毛內 輸送), 4-基底體, 5-鞭毛의 斷面, 6-基底體의 微細小管의 三重體
 軸索 의 斷面
軸索 의 斷面
 Chlamydomonas reinhardtii
의 鞭毛 領域을 通한 縱斷面. 細胞 頂點에는 鞭毛의 固定 部位인 基底部가 있다. 基底體는 9個의 周邊 微細小管 三重項이 있는 中心小體에서 由來하고 類似한 下部 構造를 가지고 있다(이미지 下段 中央의 救助 參照).
Chlamydomonas reinhardtii
의 鞭毛 領域을 通한 縱斷面. 細胞 頂點에는 鞭毛의 固定 部位인 基底部가 있다. 基底體는 9個의 周邊 微細小管 三重項이 있는 中心小體에서 由來하고 類似한 下部 構造를 가지고 있다(이미지 下段 中央의 救助 參照).
 "9+2" 救助는 axoneme의 이 橫斷面 顯微鏡 寫眞에서 볼 수 있다.
"9+2" 救助는 axoneme의 이 橫斷面 顯微鏡 寫眞에서 볼 수 있다.
이름
[
編輯
]
細菌性 偏母와 眞核生物의 纖毛 및 鞭毛의 區別을 强調하기 위해 一部 著者들은 이 두 個의 眞核生物 構造의 이름을 "undulipodia"(예: 1970年代 以後
魔窟리스
의 모든 論文)
[59]
또는 "纖毛"로 代替하려고 試圖했다. 둘 다에 對해(예: Hulsmann, 1992;
[60]
Adl et al., 2012;
[61]
캐빌리어스미스
의 大部分의 論文), 박테리아 構造에 對해 "偏母"를 保存한다. 그러나 이 記事에서 採擇된 眞核生物에 對해 "纖毛"와 "偏母"라는 用語의 差別的인 使用은 如前히 一般的이다(예: Andersen et al., 1991;
[62]
Leadbeater et al., 2000).
[63]
內部 構造
[
編輯
]
axoneme으로 알려진 眞核生物 鞭毛의 核心은 2個의 中央 單一 微細小管(單
中船
)을 둘러싸고 있는
二重線
으로 알려진 9個의 融合된
微細小管
다발이다. 이 9+2 逐朔은 眞核生物 鞭毛의 特徵이다. 眞核生物 鞭毛의 基底에는 基底體(basal body), "안檢體(blepharoplast)" 또는 키네토솜이 있으며, 이는 鞭毛 微細小管의 微細小管 組織化 中心이며 길이가 約 500나노미터이다. 基礎體는
中心小體
와 構造的으로 同一하다. 鞭毛는 細胞의
原形質膜
으로 둘러싸여 있어 鞭毛의 內部는 細胞의
細胞質
에 接近할 수 있다.
逐朔과 基底體 外에 形態가 比較的 일정하며 鞭毛 裝置의 다른 內部 構造는 전이대(軸色滯와 基底體가 만나는 곳)와 뿌리界(基底體에서 細胞質로 擴張되는 微細官 또는 原纖維 構造)이다), 眞核生物의 系統發生的 關係의 指標로서 더 다양하고 有用하다. 더 흔하지 않은 다른 構造로는 鞭毛周圍(또는 根軸, paraxonemal) 杆狀體, R 纖維 및 S 纖維가 있다.
[64]
:63?84
더 仔細한 表面 構造는 아래를 參照하라.
器具
[
編輯
]
外部 9個의 二重線 微細小管 各各은 한 雙의
디네인
인 팔("內部" 및 "外部" 팔)을 隣接한 微細小管으로 擴張한다. 이들은 ATP 加水分解를 통해 힘을 生成한다. 偏母 逐朔은 또한 放射狀 스포크, 外部 9個의 微細小管 二重線 各各에서 中央 雙을 向해 擴張되는 폴리펩타이드 複合體를 包含하며, 스포크의 "머리"는 안쪽을 向한다. 放射形 스포크는 鞭毛 運動의 調節에 關與하는 것으로 생각되지만 正確한 機能과 作用 方法은 아직 알려져 있지 않다.
[65]
偏母와 纖毛
[
編輯
]
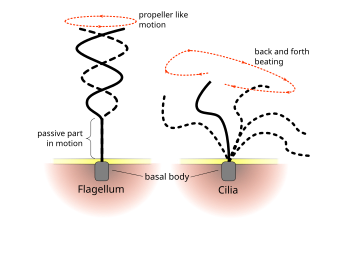 眞核生物 "flagellum"과 "cillum"의 搏動 패턴은 두 構造 以前의 傳統的인 區分으로 알려져 있다.
眞核生物 "flagellum"과 "cillum"의 搏動 패턴은 두 構造 以前의 傳統的인 區分으로 알려져 있다.
眞核生物의 纖毛와 鞭毛의 規則的인 搏動 패턴은 細胞 水準에서 움직임을 生成한다. 例를 들면
亭子
의 游泳과 같은 單一 細胞의 推進에서부터 呼吸器와 같은 細胞의 固定된 層을 따라 流體의 移動에 이르기까지 다양하다.
[66]
진핵 生物의 纖毛와 鞭毛는 窮極的으로 同一하지만 때때로 構造가 알려지기 以前의 傳統인 運動 패턴에 따라 分類된다. 鞭毛의 境遇 움직임이 平面的이고 물결 模樣인 境遇가 많지만 運動性 纖毛는 힘과 回復 스트로크로보다 複雜한 3次元 움직임을 遂行하는 境遇가 많다.
[66]
또 다른 傳統的인 區別 形態는 細胞에 있는 9+2 小器官의 數이다.
[65]
偏母 內 輸送
[
編輯
]
軸索 小段位, 幕橫斷 受容體 및 其他 蛋白質이 鞭毛의 길이를 따라 위아래로 移動하는 過程인 鞭毛 內 輸送은 運動成果 信號 傳達 모두에서 鞭毛의 適切한 機能에 必須的이다.
[67]
進化와 發生
[
編輯
]
眞核生物의 鞭毛 또는 纖毛(아마도 祖上의 特性)는
[68]
比較的 多年生 狀態로, 또는 鞭毛가 있는 生活 週期 段階(예: 조이드,
配偶者
, 有性 胞子, 持續的으로 生成되거나 아니다).
[69]
[70]
[61]
첫 番째 狀況은 "偏母 狀態"(또는 "모나도이드
組織 水準
")를 가진
纖毛
및 많은 眞核生物에서와 같이 多細胞 有機體의 特殊化된 細胞(예:
海面
의 choanocytes 또는
後生動物
의 纖毛
上皮
)에서 發見된다.
鞭毛蟲類
, 人工 그룹).
鞭毛된 壽命 週期 段階는 많은
綠藻類
(動物 胞子 및 수컷 配偶者),
蘚苔植物
(수컷 配偶者),
翼龍植物
(수컷 配偶者), 一部
겉씨植物
(
蘇鐵
및은
行
, 수컷 配偶者), 中心
閨藻類
(수컷 配偶者)와 같은 많은 그룹에서 發見된다.),
褐藻類
(動物 胞子 및 配偶者),
난균류
(無性 有性 胞子 및 配偶者),
逆帽菌類
(動物 胞子),
亡兄蟲類
(動物 胞子), 一部
頂端複合體蟲類
(配偶者), 一部
放散蟲
(아마 배우자),
[71]
有孔蟲
(
配偶者
) (動物 胞子 및 配偶者),
變形菌江
(動物 胞子),
動物
(男性 生殖 細胞) 및
護喪菌類
(動物 胞子와 配偶者).
偏母 또는 纖毛는 原始 狀態가 아니라 損失로 인해 一部 그룹에서 完全히 不在한다. 纖毛의 小室은
紅藻類
, 一部 綠藻類(
接合鳥綱
), 소철류와 銀杏나무를 除外한
겉씨植物
,
속씨植物
, 5開形
돌末流
, 一部
頂端複合體蟲類
, 一部
아메바類
, 一部
後生動物
의 亭子,
[72]
및
眞菌類
(
除外
).
類型學
[
編輯
]
偏母 또는 纖毛와 關聯된 많은 用語가 眞核生物을 특징짓는 데 使用된다.
[70]
[73]
[64]
:60?63
[74]
[75]
存在하는 表面 構造에 따라 鞭毛는 다음과 같을 수 있다.
- 偏母 鞭毛(= 매끄럽고 末端 鞭毛): 털이 없음, 例를 들어
後偏母生物
에서
- 털이 많은 偏母(= tinsel, flimmer, pleuronematic 鞭毛): 털이 있는(= mastigonemes
sensu lato
), 다음으로 나뉜다.
- 스티코네마틱 鞭毛: 한 줄의 털이 있는 偏母
- 版吐네틱 鞭毛: 두 줄의 털이 있음
- acronematic: 單一 末端 유선모 또는 鞭毛를 가진 鞭毛(예: bodonids);
[76]
- 비늘 包含: 예: Prasinophyceae
- 가시가 있는 境遇: 예: 一部
褐藻類
- 물결 模樣의 膜이 있는 境遇: 例를 들어 一部 키네토플라스티드, 一部
浮氣滯留
,
運動核鞭毛蟲類
等이다.
- 코와 함께(細胞의 줄기와 같은 突出部): 예:
阿푸塑造아
, 一部 bodonids
[77]
鞭毛의 數에 따라 細胞는 다음과 같을 수 있다. (一部 著者는 "flagellated" 代身 "ciliated"를 使用함을 記憶하라)
[61]
[78]
- unflagellated: 예, 大部分의
Opisthokonta
- biflagellated
: 예, 모든
Dinoflagellata
,
Charophyceae
의 配偶者, 大部分의 蘚苔 動物 및 一部
後生 動物
[72]
- 三鞭毛: 예, 一部
有孔蟲
의 配偶者
- 四面像: 예, 一部 Prasinophyceae,
Collodictyonidae
- octoflagellated: 예, 一部
Diplomonada
, 一部 Prasinophyceae
- 多重 鞭毛: 예:
Opalinata
, Ciliophora,
Stephanopogon
,
Parabasalida
, Hemimastigophora,
Caryoblastea
,
Multicilia
,
Oedogoniales
(
Chlorophyta
)의 配偶者 (또는 조이드), 一部
多核아베마속
과
겉씨植物
과
羊齒植物
.
鞭毛의 揷入 位置에 따라:
[79]
- opisthokont
: Opisthokonta에서와 같이 뒤쪽에 鞭毛가 揷入된 細胞(Vischer, 1945).
Haptophyceae
에서 鞭毛는 側面에서 末端으로 揷入되지만 빠른 水泳 中에는 뒤쪽으로 向한다.
[80]
- akrokont: 鞭毛가 頂點에 揷入된 細胞
- 아크로콘트(subakrokont): 鞭毛가 阿諂에 揷入된 細胞
- pleurokont: 鞭毛가 옆으로 揷入된 細胞
搏動 패턴에 따르면:
- 滑空: 氣質을 따라가는 鞭毛
[77]
- heterodynamic: 다른 搏動
[81]
을 가진 鞭毛
- isodynamic: 같은 패턴의 鞭毛 搏動
偏母 類型과 關聯된 기타 用語:
古筠
[
編輯
]
一部
古筠
種의 鞭毛는 細菌의 偏母와 表面的으로 類似하다. 1980年代에는 全體的인 形態와 行動에 기초하여 둘의 鞭毛가 上洞人 것으로 생각되었다. 偏母와 古筠은 모두 細胞 外部로 뻗어 있는 필라멘트로 構成되어 있으며 回轉하여 細胞를 推進한다. Archaeal 鞭毛는 中央 채널이 없는 獨特한 構造를 가지고 있다. 박테리아
類型 IV 筆린
과 類似하게, 古筠 蛋白質(아르케엘린)은 클래스 3 信號 펩티드로 만들어지며 類型 IV 프리筆린 펩티다제 類似 酵素에 依해 處理된다. 아르케엘린은 一般的으로 適切한 組立 또는 機能에 必要한 N-連結
글리칸
의 追加에 依해 變形된다.
[3]
1990年代의 發見은 古筠과 細菌 鞭毛 사이의 수많은 細部的인 差異點을 밝혀냈다. 여기에는 다음이 包含된다.
- 細菌의 鞭毛 回轉은
H
+
이온
의 흐름인
陽性子 驅動力
또는 때로는
Na
+
이온의 흐름인 나트륨 原動力에 依해 구동된다. 古細菌 鞭毛 回轉은
ATP
에 依해 구동된다.
[82]
- 박테리아 細胞에는 各各 獨立的으로 回轉하는 많은 偏母 필라멘트가 있는 境遇가 많지만, 古細菌 鞭毛는 單一 어셈블리로 回轉하는 많은 필라멘트 다발로 構成된다.
- 細菌 鞭毛는 끝에 鞭毛 小段位가 追加되어 자란다. 古筠 鞭毛는 基底部에 小段位體를 追加함으로써 자란다.
- 細菌性 鞭毛는 古筠보다 두껍고 細菌性 필라멘트 內部에는 鞭毛 小單位體가 필라멘트의 內部로 흘러들어올 수 있고 끝에 追加될 수 있는 充分히 큰 속이 빈 "棺"李 內部에 있다.
[83]
- 細菌性 鞭毛의 많은 構成 要素는
III型 分泌 시스템
의 構成 要素와 序列 類似性을 共有하지만 細菌性 偏母와 古細菌의 構成 要素는 序列 類似性을 共有하지 않는다. 代身, 古細菌의 一部 構成 要素는
II型 分泌 시스템
의 作用을 통해 組立되는
類型 IV
필리의 構成 要素와 序列 및 形態學的 類似性을 共有한다(단백질 分泌 시스템의 命名法은 一貫되지 않음).
[83]
이러한 差異點은 세균의 偏母와 古細菌이
相同性
이라기보다는 生物學的
類推
또는
收斂 鎭火
의 古典的 事例가 될 수 있음을 의미한다.
[84]
[85]
그러나 數十 年 동안 잘 알려진 박테리아 鞭毛 硏究(예:Howard Berg)와 比較할 때,
[86]
古細菌은 最近에야 과학적 關心을 받기 始作했다.
追加 이미지
[
編輯
]
各州
[
編輯
]
- ↑
“Prokaryotic motility structures”. 《Microbiology》
149
(Pt 2): 295?304. February 2003.
doi
:
10.1099/mic.0.25948-0
.
PMID
12624192
.
- ↑
“Assembly and motility of eukaryotic cilia and flagella. Lessons from Chlamydomonas reinhardtii”
. 《Plant Physiology》
127
(4): 1500?7. December 2001.
doi
:
10.1104/pp.010807
.
PMC
1540183
.
PMID
11743094
.
- ↑
가
나
《Pili and Flagella: Current Research and Future Trends》. Caister Academic Press. 2009.
ISBN
978-1-904455-48-6
.
- ↑
“Sperm design and sperm function”
. 《Biology Letters》
2
(2): 246?9. June 2006.
doi
:
10.1098/rsbl.2006.0449
.
PMC
1618917
.
PMID
17148374
.
- ↑
“Helicobacter pylori: ulcers and more: the beginning of an era”
. 《The Journal of Nutrition》
131
(10): 2789S?2793S. October 2001.
doi
:
10.1093/jn/131.10.2789S
.
PMID
11584108
. 2009年 2月 7日에
原本 文書
(abstract page)
에서 保存된 文書
. 2008年 6月 2日에 確認함
.
- ↑
“Sensing wetness: a new role for the bacterial flagellum”
. 《The EMBO Journal》
24
(11): 2034?42. June 2005.
doi
:
10.1038/sj.emboj.7600668
.
PMC
1142604
.
PMID
15889148
.
- ↑
“The archaellum: how Archaea swim”
. 《Frontiers in Microbiology》
6
: 23. 2015年 1月 27日.
doi
:
10.3389/fmicb.2015.00023
.
PMC
4307647
.
PMID
25699024
.
- ↑
Quax, TEF; Albers, SV; Pfeiffer, F (2018年 12月 14日).
“Taxis in archaea.”
. 《Emerging Topics in Life Sciences》
2
(4): 535?546.
doi
:
10.1042/ETLS20180089
.
PMC
7289035
.
PMID
33525831
.
- ↑
“Cilia, flagella, and microtubules”
. 《The Journal of Cell Biology》
91
(3 Pt 2): 125s?130s. December 1981.
doi
:
10.1083/jcb.91.3.125s
.
PMC
2112827
.
PMID
6459327
.
- ↑
“Flagellar rotation in the archaeon Halobacterium salinarum depends on ATP”. 《Journal of Molecular Biology》
384
(1): 1?8. 2008.
doi
:
10.1016/j.jmb.2008.08.057
.
PMID
18786541
.
- ↑
가
나
Alberts, Bruce (2015). 《Molecular biology of the cell》 Six版. New York, NY. 942쪽.
ISBN
9780815344643
.
- ↑
“Flagellar rotation and the mechanism of bacterial motility”. 《Nature》
249
(452): 73?4. May 1974.
Bibcode
:
1974Natur.249...73S
.
doi
:
10.1038/249073a0
.
PMID
4598030
.
- ↑
“Rapid rotation of flagellar bundles in swimming bacteria”. 《Nature》
325
(6105): 637?640. 1987.
Bibcode
:
1987Natur.325..637L
.
doi
:
10.1038/325637a0
.
- ↑
“Bacteria swim by rotating their flagellar filaments”. 《Nature》
245
(5425): 380?2. October 1973.
Bibcode
:
1973Natur.245..380B
.
doi
:
10.1038/245380a0
.
PMID
4593496
.
- ↑
“Movement and locomotion of microorganisms”. 《Annual Review of Microbiology》
19
: 21?58. 1965.
doi
:
10.1146/annurev.mi.19.100165.000321
.
PMID
5318439
.
- ↑
“Bacterial motility on a surface: many ways to a common goal”. 《Annual Review of Microbiology》
57
: 249?73. 2003.
doi
:
10.1146/annurev.micro.57.030502.091014
.
PMID
14527279
.
- ↑
“Archaeal flagella, bacterial flagella and type IV pili: a comparison of genes and posttranslational modifications”. 《Journal of Molecular Microbiology and Biotechnology》
11
(3?5): 167?91. 2006.
doi
:
10.1159/000094053
.
PMID
16983194
.
- ↑
“Bacterial and archaeal flagella as prokaryotic motility organelles”. 《Biochemistry. Biokhimiia》
69
(11): 1203?12. November 2004.
doi
:
10.1007/s10541-005-0065-8
.
PMID
15627373
.
- ↑
〈Archaeal Flagella and Pili〉. 《Pili and Flagella: Current Research and Future Trends》. Caister Academic Press. 2009.
ISBN
978-1-904455-48-6
.
- ↑
“How bacteria assemble flagella”. 《Annual Review of Microbiology》
57
: 77?100. 2003.
doi
:
10.1146/annurev.micro.57.030502.090832
.
PMID
12730325
.
- ↑
“Stabilization of flagellar filaments by HAP2 capping”. 《FEBS Letters》
568
(1?3): 105?9. June 2004.
doi
:
10.1016/j.febslet.2004.05.029
.
PMID
15196929
.
- ↑
“Divergence of quaternary structures among bacterial flagellar filaments”. 《Science》
320
(5874): 382?5. April 2008.
Bibcode
:
2008Sci...320..382G
.
doi
:
10.1126/science.1155307
.
PMID
18420936
.
- ↑
“Polar and lateral flagellar motors of marine Vibrio are driven by different ion-motive forces”. 《Nature》
355
(6356): 182?4. January 1992.
Bibcode
:
1992Natur.355..182A
.
doi
:
10.1038/355182a0
.
PMID
1309599
.
- ↑
Dean, Tim (2010年 8月 2日).
“Inside nature's most efficient motor: the flagellar”
. 《Australian Life Scientist》.
- ↑
Nagata, Yoshio (June 2014).
“Unlocking the secrets of nature's nanomotor”
. 《Nikkei Asian Review》.
- ↑
“Steps in the bacterial flagellar motor”
. 《PLOS Computational Biology》
5
(10): e1000540. October 2009.
arXiv
:
0904.0438
.
Bibcode
:
2009PLSCB...5E0540M
.
doi
:
10.1371/journal.pcbi.1000540
.
PMC
2759076
.
PMID
19851449
.
- ↑
Whitfield, John (2008年 6月 19日).
“Bacterial engines have their own clutch”
. 《Nature News》 (英語): news.2008.903.
doi
:
10.1038/news.2008.903
. 2017年 5月 17日에 確認함
.
- ↑
〈Chapter 13〉. 《Living at Micro Scale: The Unexpected Physics of Being Small》. Cambridge: Harvard University Press. 2009.
ISBN
978-0-674-03116-6
.
- ↑
Hildebrand, Milton (November 1959). “Motions of the running Cheetah and Horse”. 《
Journal of Mammalogy
》
44
(4): 481?495.
doi
:
10.2307/1376265
.
JSTOR
1376265
.
Although according to
Hunter, Luke; Hamman, Dave (2003). 《Cheetah》. Struik Publishers. 37?38쪽.
the cheetah's fastest recorded speed was 110 km/h (68 mph)
- ↑
“How bacteria shift gears”
. 《PLOS Biology》
9
(5): e1001061. May 2011.
doi
:
10.1371/journal.pbio.1001061
.
PMC
3091840
.
PMID
21572986
.
- ↑
“Mechanisms of type III protein export for bacterial flagellar assembly”. 《Molecular BioSystems》
4
(11): 1105?15. November 2008.
doi
:
10.1039/b808065h
.
PMID
18931786
.
- ↑
“Reconstitution of Bacterial Flagella In Vitro”. 《Journal of Molecular Biology》
10
: 42?56. October 1964.
doi
:
10.1016/S0022-2836(64)80026-7
.
PMID
14222895
.
- ↑
가
나
“From The Origin of Species to the origin of bacterial flagella”. 《Nature Reviews. Microbiology》
4
(10): 784?90. October 2006.
doi
:
10.1038/nrmicro1493
.
PMID
16953248
.
- ↑
가
나
“Evolution of bacterial type III protein secretion systems”. 《Trends in Microbiology》
12
(3): 113?5. March 2004.
doi
:
10.1016/j.tim.2004.01.003
.
PMID
15001186
.
- ↑
“Bacterial type III secretion systems are ancient and evolved by multiple horizontal-transfer events”. 《Gene》
312
: 151?63. July 2003.
doi
:
10.1016/S0378-1119(03)00612-7
.
PMID
12909351
.
- ↑
“Evolution of the type III secretion system and its effectors in plant-microbe interactions”. 《The New Phytologist》
177
(1): 33?47. 2008.
doi
:
10.1111/J.1469-8137.2007.02293.X
.
PMID
18078471
.
- ↑
Behe, M. (2007) The Edge of Evolution. Free Press, New York
- ↑
“The protein network of bacterial motility”
. 《Molecular Systems Biology》
3
: 128. 2007.
doi
:
10.1038/msb4100166
.
PMC
1943423
.
PMID
17667950
.
- ↑
“Novel conserved assembly factor of the bacterial flagellum”
. 《Journal of Bacteriology》
188
(21): 7700?6. November 2006.
doi
:
10.1128/JB.00820-06
.
PMC
1636259
.
PMID
16936039
.
- ↑
“Novel genes associated with enhanced motility of Escherichia coli ST131”
. 《PLOS ONE》
12
(5): e0176290. 2017年 5月 10日.
Bibcode
:
2017PLoSO..1276290K
.
doi
:
10.1371/journal.pone.0176290
.
PMC
5425062
.
PMID
28489862
.
- ↑
“Bacterial flagella and Type III secretion: case studies in the evolution of complexity”. 《Genome Dynamics》
3
: 30?47. 2005.
doi
:
10.1159/000107602
.
ISBN
978-3-8055-8340-4
.
PMID
18753783
.
- ↑
“Bacterial flagella”
(PDF)
. 2021年 12月 29日에 確認함
.
- ↑
“Architecture of a flagellar apparatus in the fast-swimming magnetotactic bacterium MO-1”
. 《Proc Natl Acad Sci U S A》
109
(50): 20643?8. December 2012.
Bibcode
:
2012PNAS..10920643R
.
doi
:
10.1073/pnas.1215274109
.
PMC
3528567
.
PMID
23184985
.
- ↑
“tricho- prefix”
. 2022年 3月 26日에 確認함
.
- ↑
Echazarreta, MA; Klose, KE (2019).
“
Vibrio
Flagellar Synthesis.”
. 《Frontiers in Cellular and Infection Microbiology》
9
: 131.
doi
:
10.3389/fcimb.2019.00131
.
PMC
6504787
.
PMID
31119103
.
- ↑
“Lopho”
. 2022年 3月 26日에 確認함
.
- ↑
“Cryo-electron tomography elucidates the molecular architecture of Treponema pallidum, the syphilis spirochete”
. 《Journal of Bacteriology》
191
(24): 7566?80. December 2009.
doi
:
10.1128/JB.01031-09
.
PMC
2786590
.
PMID
19820083
.
- ↑
“Native cellular architecture of Treponema denticola revealed by cryo-electron tomography”
. 《Journal of Structural Biology》
163
(1): 10?7. July 2008.
doi
:
10.1016/j.jsb.2008.03.009
.
PMC
2519799
.
PMID
18468917
.
- ↑
“Comparative cryo-electron tomography of pathogenic Lyme disease spirochetes”. 《Molecular Microbiology》
71
(6): 1415?34. March 2009.
doi
:
10.1111/j.1365-2958.2009.06613.x
.
PMID
19210619
.
- ↑
“Analysis of the polar flagellar gene system of Vibrio parahaemolyticus”
. 《Journal of Bacteriology》
182
(13): 3693?704. July 2000.
doi
:
10.1128/JB.182.13.3693-3704.2000
.
PMC
94540
.
PMID
10850984
.
- ↑
“Effect of viscosity on swimming by the lateral and polar flagella of Vibrio alginolyticus”
. 《Journal of Bacteriology》
178
(16): 5024?6. August 1996.
doi
:
10.1128/jb.178.16.5024-5026.1996
.
PMC
178290
.
PMID
8759871
.
- ↑
“Dual flagellar systems enable motility under different circumstances”. 《Journal of Molecular Microbiology and Biotechnology》
7
(1?2): 18?29. 2004.
doi
:
10.1159/000077866
.
PMID
15170400
.
- ↑
“Bacterial lateral flagella: an inducible flagella system”. 《FEMS Microbiology Letters》
263
(2): 127?35. October 2006.
doi
:
10.1111/j.1574-6968.2006.00403.x
.
PMID
16978346
.
- ↑
“Regulation of lateral flagella gene transcription in Vibrio parahaemolyticus”
. 《Journal of Bacteriology》
167
(1): 210?8. July 1986.
doi
:
10.1128/jb.167.1.210-218.1986
.
PMC
212863
.
PMID
3013835
.
- ↑
“Analysis of the lateral flagellar gene system of Aeromonas hydrophila AH-3”
. 《Journal of Bacteriology》
188
(3): 852?62. February 2006.
doi
:
10.1128/JB.188.3.852-862.2006
.
PMC
1347325
.
PMID
16428388
.
- ↑
“Polar flagellum biogenesis in Aeromonas hydrophila”
. 《Journal of Bacteriology》
188
(2): 542?55. January 2006.
doi
:
10.1128/JB.188.2.542-555.2006
.
PMC
1347287
.
PMID
16385045
.
- ↑
“A macroscopic scale model of bacterial flagellar bundling”
. 《Proceedings of the National Academy of Sciences of the United States of America》
100
(26): 15481?5. December 2003.
arXiv
:
cond-mat/0312562
.
Bibcode
:
2003PNAS..10015481K
.
doi
:
10.1073/pnas.2633596100
.
PMC
307593
.
PMID
14671319
.
- ↑
“Bacterial flagella rotating in bundles: a study in helical geometry”
. 《Proceedings of the National Academy of Sciences of the United States of America》
74
(1): 221?5. January 1977.
Bibcode
:
1977PNAS...74..221M
.
doi
:
10.1073/pnas.74.1.221
.
PMC
393230
.
PMID
264676
.
- ↑
“The collapse of the two-kingdom system, the rise of protistology and the founding of the International Society for Evolutionary Protistology (ISEP)”. 《International Journal of Systematic and Evolutionary Microbiology》
53
(Pt 6): 1707?14. November 2003.
doi
:
10.1099/ijs.0.02587-0
.
PMID
14657097
.
- ↑
“Undulipodium: End of a useless discussion”. 《European Journal of Protistology》
28
(3): 253?7. August 1992.
doi
:
10.1016/s0932-4739(11)80231-2
.
PMID
23195228
.
- ↑
가
나
다
“The revised classification of eukaryotes”
. 《The Journal of Eukaryotic Microbiology》
59
(5): 429?93. September 2012.
doi
:
10.1111/j.1550-7408.2012.00644.x
.
PMC
3483872
.
PMID
23020233
.
- ↑
“Terminology and nomenclature of the cytoskeletal elements associated with the flagellar/ciliary apparatus in protists”. 《Protoplasma》
164
(1?3): 1?8. 1991.
doi
:
10.1007/bf01320809
.
- ↑
Leadbeater, Barry S. C.; Green, John C., 編輯. (2000).
《Flagellates: Unity, Diversity and Evolution》
. The Systematics Association Special Volume
59
. Taylor and Francis.
ISBN
978-1-4822-6822-5
.
- ↑
가
나
Barsanti, Laura; Gualtieri, Paolo (2006).
《Algae: Anatomy, Biochemistry, and Biotechnology》
. Florida, USA: CRC Press.
ISBN
9780203492598
.
- ↑
가
나
Lindemann, CB; Lesich, KA (2010年 2月 15日). “Flagellar and ciliary beating: the proven and the possible.”. 《Journal of Cell Science》
123
(Pt 4): 519?28.
doi
:
10.1242/jcs.051326
.
PMID
20145000
.
- ↑
가
나
Lodish, Harvey; Berk, Arnold; Zipursky, S. Lawrence; Matsudaira, Paul; Baltimore, David; Darnell, James (2000). 〈Section 19.4Cilia and Flagella: Structure and Movement〉.
《Cilia and Flagella: Structure and Movement》
.
ISBN
0-7167-3136-3
.
- ↑
“Intraflagellar transport and cilia-dependent renal disease: the ciliary hypothesis of polycystic kidney disease”. 《Journal of the American Society of Nephrology》
15
(10): 2528?36. October 2004.
doi
:
10.1097/01.ASN.0000141055.57643.E0
.
PMID
15466257
.
- ↑
“Evolution of microtubule organizing centers across the tree of eukaryotes”. 《The Plant Journal》
75
(2): 230?44. July 2013.
doi
:
10.1111/tpj.12145
.
PMID
23398214
.
- ↑
Raven, J.A. (2000). 〈The flagellate condition〉. 《
Leadbeater & Green 2000
, 27?48쪽
括弧 없는 하버드 認容 error: 對象 없음: CITEREFLeadbeaterGreen2000 (
help
)
》.
ISBN
9781482268225
.
- ↑
가
나
〈Spores of Fungi〉. 《2007》 3板. Cambridge: Cambridge University Press. 2007年 1月 25日. 23?24쪽.
ISBN
9781139461504
.
- ↑
“The chastity of amoebae: re-evaluating evidence for sex in amoeboid organisms”
. 《Proceedings. Biological Sciences》
278
(1715): 2081?90. July 2011.
doi
:
10.1098/rspb.2011.0289
.
PMC
3107637
.
PMID
21429931
.
- ↑
가
나
〈Evolution of human gametes: spermatozoa.〉. 《Gametes: the spermatozoon》. Cambridge University Press. 1995.
ISBN
9780521479967
.
- ↑
《Introduction to Phycology》
. Oxford: Blackwell Scientific Publications. 1987. 65쪽.
ISBN
9781444314205
.
- ↑
《The Fine Structure of Algal Cells》
. London: Academic Press. 1973. 57?79쪽.
ISBN
9780323158237
.
- ↑
《Phycology》
4板. Cambridge University Press. 2008.
7
쪽.
ISBN
9781139469876
.
lee tubular hairs.
- ↑
Corliss, J.O.; Lom, J (2000). 〈An annotated glossary of protozoological terms〉. Lee, J.J.; Leedale, G.F.; Bradbury, P. 《An illustrated guide to the protozoa》
2
2板. Society of Protozoologists. 1346?85쪽.
ISBN
1891276239
.
- ↑
가
나
“A short guide to common heterotrophic flagellates of freshwater habitats based on the morphology of living organisms”. 《Protist》
164
(6): 842?60. November 2013.
doi
:
10.1016/j.protis.2013.08.003
.
PMID
24239731
.
- ↑
《Protozoa and other Protists》
. London: Edward Arnold. 1989. 98?99쪽.
ISBN
9780521428057
.
- ↑
《Aquatic phycomycetes》
2板. Ann Arbor: Michigan: University of Michigan Press. 1960.
15
쪽.
- ↑
“The ultrastructure and taxonomy of the Chrysophyceae and Prymnesiophyceae (Haptophyceae): a survey with some new observations on the ultrastructure of the Chrysophyceae”. 《Journal of the Linnean Society of London, Botany》
72
(2): 55?80. 1976.
doi
:
10.1111/j.1095-8339.1976.tb01352.x
.
- ↑
“Origin and evolution of flagellar movement”
. 《Cell Motil》
5
: 137?138. 1985.
- ↑
Madigan, Michael T. (2019). 《Brock biology of microorganisms》 Fifteen, Global版. NY, NY. 70?71쪽.
ISBN
9781292235103
.
- ↑
가
나
“Assembly and function of the archaeal flagellum”. 《Biochemical Society Transactions》
39
(1): 64?9. January 2011.
doi
:
10.1042/BST0390064
.
PMID
21265748
.
- ↑
“The archaeal flagellum: a different kind of prokaryotic motility structure”. 《FEMS Microbiology Reviews》
25
(2): 147?74. April 2001.
doi
:
10.1111/j.1574-6976.2001.tb00575.x
.
PMID
11250034
.
- ↑
“Archaeal flagellum”
. 《www.uniprot.org》
. 2019年 6月 24日에 確認함
.
- ↑
Berg, Howard C. (2003). 《E. coli in motion》 1. Aufl.판. New York: Springer.
ISBN
9780387008882
.