古細菌
(古細菌,
라틴語
:
Archaea
아르케아
[
*
]
) 또는
古筠
(古菌)은
單細胞
生物
分類群의 하나이다. 細胞核이 없는 微生物로
原核生物
의 한 部類이며, 細菌과 다른 系를 이루고 있다는 것이 最近에 밝혀졌다.
細菌
,
眞核生物
과 함께 3個의 生物
驛
中 하나이다.
[1]
어떤 點은
細菌
과 닮아있고, 어떤 點은
眞核生物
과 닮아있다.
細菌과의 比較
[
編輯
]
- 共通點
古細菌과
細菌
은 共通的으로
진핵細胞
에서 볼 수 있는 核膜이나 膜性小器官이 없고 긴 圓形의
染色體
DNA를 갖고 있으며 間或 작은
플라스미드
를 갖고 있는 境遇도 있다. 또한 特定
大使
와 聯關된 古細菌과 細菌의
遺傳子
는 共히
오페론
을 이루거나 群集을 이루고 있는 境遇가 많으며 古細菌과 細菌 모두 제2형 制限 酵素(認識部位와 切斷部位가 모두 特異性이 있는 制限酵素) 시스템을 가진다.
- 差異點
하지만 古細菌과
細菌
을 가르는 가장 큰 差異點은 크게 세 가지 程度로 要約할 수 있다. 첫째는
遺傳子
및
蛋白質
의 相同性, 둘째는
細胞壁
과
細胞膜
의 構成, 셋째는
DNA
複製 및
蛋白質 合成
에서의
진핵細胞
와의 類似性이다. 古細菌은 細胞膜을 構成하는
燐脂質
의 構造에 있어서도 細菌과 約 4가지의 差異點을 보이는데, 첫째 글리세롤의 方向性(L-form), 둘째
에테르
結合, 셋째
이소프레盧이드
사슬, 넷째, 가지가 있다는 點이다. 普通 細菌과 幕 構成成分이 다르기 때문에
抗生劑
로도 破壞되지 않는다. 細胞壁의 境遇에도 細菌은
펩티도글리칸
層을, 古細菌은
슈道펩티도글리칸
層을 갖는다.
古細菌의 種類
[
編輯
]
特徵과 棲息地에 따라 區分되지만 여러 가지 特徵을 同時에 지닌 古細菌度 있다. 基本的으로 메탄生成菌, 極好鹽性菌, 好熱性菌, 超高溫性菌으로 區分한다.
메테人菌
[
編輯
]
메테人 細菌
은 CO
2
와 H
2
를
메테人
가스(CH
4
)로 轉換할 수 있다. 이 過程을 메테人生成이라고 부른다. 메테人 細菌은 흔히 無酸素性
진흙
이나 江과 바다 바닥의 沈澱物에서 살지만,
口腔
이나
大將
에서 사는 種도 있다.
[2]
生理學的으로 메탄生成細菌은 絶對 嫌氣性菌으로 그들을 培養하기 위해 絶對 嫌氣性 技術이 必要하다. 種들에 따라서는 메탄生成細菌의 培養은 CO
2
와 H
2
의 空氣로 된 最小배지에서 可能하다. 비록
極限環境 微生物
종들은 매우 높거나 낮은 溫度 또는 鹽의 濃度가 매우 높은 곳에서 最適으로 成長하지만 大部分의 알려진 메탄生成細菌들은 重穩城과 非好鹽性이다.
極好鹽性 細菌
[
編輯
]
極好鹽性 細菌들은 天日鹽田, 天然 鹽湖水, 或은 人工的인 鹽分 棲息地와 相當히 많은 소금으로 절인 魚類와 肉類의 表面과 같은 매우 많은 鹽分이 있는 環境에 사는 다양한 그룹이다. 極好鹽性 細菌은 成長을 위하여 적어도 1.5M(藥 9%) NaCl이 必要한 生物이다. 大部分의 鐘들은 適正 成長을 위하여 2~4M NaCl(12~23%)을 必要로 한다. 事實上 모든 極好鹽性 細菌들은 비록 몇몇 종들이 이 濃度에서 매우 느리게 成長하지만, 5.5M NaCl(32%, NaCl의 飽和 限度)에서 成長할 수 있다.
極好鹽性 細菌들은 그람音聲이며 二分法에 依해 增殖하고 休眠段階나 胞子를 形成하지 않는다. 大部分의 好鹽性菌은 運動性이 없으나 몇 種類에서는 鞭毛를 가지고 徐徐히 움직이거나 가스 小包(gas vesicles)로 떠다닐 수 있다.
黠虜朴테리움
과 黠虜콕쿠스의 遺傳體 조성은 全體 細胞 DNA의 25~30% 以上을 包含하는 큰 플라스미드들이 存在하고 이들 플라스미드들의 GC鹽基 比率(57~60% GC)李 染色體 DNA의 것(66~68%)과 매우 다르다는 點에서 特異하다. 極好鹽性 細菌에서 由來하는 플라스미드는 自然系에서 알려진 가장 큰 플라스미드 中의 하나이다.
極好鹽性 細菌의 大部分의 鐘들은 絶對 好氣性이다. 大部分 好鹽性菌은 아미노산 或은 有機酸을 에너지源으로 使用하고 最適의 成長을 위하여 많은 種類의 成長因子(主로 비타민들)를 必要로 한다. 어떤 黠虜朴테리움 종들은 炭水化物을 酸化하나 이런 能力은 比較的 드물다. 黠虜朴테리움에는 시토크롬 a, b, c 兄을 包含하는 傳達系가 存在하고, 에너지는 細胞膜에 依해 媒介되는 化學渗透 現象으로 生成된 陽性子 動力을 通해서 好機的으로 成長하는 것으로 알려져 있다. 어떤 好鹽性 古細菌은 嫌忌敵으로 成長하는 것으로 알려져 있다. 黨 醱酵를 하거나 窒酸鹽 或은 푸마르산의 還元과 連結된 嫌氣性 呼吸(anaerobic respiration)에 依하여 嫌氣性腸(anoxic growth)하는 것이 몇몇 種들에서 報告된 바 있다.
好熱菌
[
編輯
]
好熱菌
은 다른 細菌에 比해 相對的으로 45 °C 以上에서도 生存한다. 好熱菌은 地球의 뜨거운 地域에서 많이 發見되는데, 肺石炭과 같이 自發的인 燃燒에 依하여 熱을 내는 地域에서 자주 發見된다. 뜨거운 肺石炭에서 流出되는 有機化合物을 大寫하고 繁殖한다.
테르모플라스마
의 두 番째 종인 T.volcanium은 全 世界에 걸쳐 高溫의 酸性 土壤에서 分離되었는데, 여러 個의 偏母에 依한 높은 運動性을 나타낸다. 細胞壁이 없이 渗透壓 스트레스에 生存하고, 낮은 pH와 높은 溫度라는 兩쪽의 極限 環境에 견디기 위해서 테르모플라스마는 獨特한 細胞膜 構造를 進化시켰다. 細胞膜은 liposaccharide이라 불리는 脂質多糖類 類似物質을 包含하고 있다. 테르모플라스마의 DNA는 높은 鹽基性의 DNA 結合蛋白質과 複合體를 이루어 求刑 粒子 속으로 DNA를 組織化하는데, 이는 眞核生物의 뉴클레오솜과 類似하다. 이 蛋白質은 眞核生物의 鹽基性 히스톤 段質과 類似性이 있다. 히스톤과 비슷한 蛋白質들이 여러 다른
에우리古細菌
에서 發見되고 있다.
超高溫性 細菌
[
編輯
]
地表面 數千미터 및, 溫度와 壓力이 매우 높은 곳에서도 繁殖하는 細菌이다. hyperthermophiles는 油類의 環境에서 SO₄²?를 黃化水素로 還元시켜 石油의 質과 回收率을 저하시키고, 海洋 石油 플랫폼을 支撐하는 鐵과 鋼鐵 合金을 腐蝕시킨다. 技術者들은 오랫동안 石油 플랫폼에서 黃化水素의 問題點을 알고 있었으나, 이것이 hyperthermophiles 때문이라는 것은 最近에 알게 되었다. 甚해 高熱 噴出口 近處에 사는 細菌은 約 2m 程度의 서관충과 함께 共生하며 살아가기도 한다. 서관충은 죽은 細菌과 細菌이 黃化物을 酸化시킬 때 放出하는 化學物質을 消費한다.
噴出孔 周邊에서 發見되는 古細菌은 바이오메탄, 바이오水素, 極限酵素 生産 等 生命工學 分野에서 다양한 素材로 活用되고 있는 單細胞 微生物을 말한다. 이들은 無酸素, 高溫, 高氣壓 等 極限의 環境에서 生存함으로써 놀라운 生命力을 보여줌과 同時에 水素, 메탄가스와 같은 代替 에너지를 만들어 내는 能力을 保有하고 있어 最近 世界的으로 많은 硏究가 進行 中이다.
또한 高溫의 열수가 存在하는 環境에서도 安定的인 蛋白質 構造를 維持하는 理由를 밝혀낼 수 있다면 産業 新素材 開發에 貴重한 硏究가 될 수 있다.
細菌, 古細菌, 眞核生物
[
編輯
]
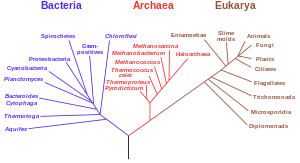 rRNA
에 基盤한
系統樹
. 細菌과 古細菌, 眞核生物의 分化를 보여준다.
rRNA
에 基盤한
系統樹
. 細菌과 古細菌, 眞核生物의 分化를 보여준다.
보라: 細菌, 빨강: 古筠, 褐色: 眞核生物
다음은 2006年
토머스 캐빌리어스미스
가 提案한 生命의 나무 系統 分類이다.
[3]
다음은 古細菌의 系統 分類이다.
[4]
[5]
[6]
| 古細菌
|
|
|
韻딘古細菌
|
|
|
|
|
|
|
後베르古細菌/아이니그므古細菌/
나노호염古細菌
/
나노古細菌
|
|
|
|
|
|
|
Altarchaeota/Diapherotrites/미크르古細菌
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
도메인 分類
[
編輯
]
| 린네
|
헤켈
|
채튼
|
코플랜드
|
휘태커
|
워즈
|
캐빌리어스미스
|
1735年
2係 分類
|
1866年
3係 分類
|
1925年
2係 分類
|
1938年
4係 分類
|
1969年
5係 分類
|
1977年
6係 分類
|
1990年
3役 分類
|
1993年
8係 分類
|
1998年
6係 分類
|
| (다루지 않음)
|
原生生物
(Protista)
|
原核生物
(Prokaryota)
|
모네라
(Monera)
|
모네라
(Monera)
|
眞正細菌
(Eubacteria)
|
細菌
(Bacteria)
|
眞正細菌
(Eubacteria)
|
細菌
(bacteria)
|
古細菌
(Archaeabacteria)
|
古細菌
(Archaea)
|
古細菌
(Archaeabacteria)
|
眞核生物
(Eukaryota)
|
原生生物
(Protista)
|
原生生物
(Protista)
|
原生生物
(Protista)
|
眞核生物
(Eukarya)
|
高動物
(Archezoa)
|
原生動物
(Protozoa)
|
原生動物
(Protozoa)
|
有色組植物
(Chromista)
|
有色組植物
(Chromista)
|
植物
(Vegetabilia)
|
植物
(Plantae)
|
植物
(Plantae)
|
植物
(Plantae)
|
植物
(Plantae)
|
植物
(Plantae)
|
植物
(Plantae)
|
菌類
(Fungi)
|
菌類
(Fungi)
|
菌類
(Fungi)
|
菌類
(Fungi)
|
動物
(Animalia)
|
動物
(Animalia)
|
動物
(Animalia)
|
動物
(Animalia)
|
動物
(Animalia)
|
動物
(Animalia)
|
動物
(Animalia)
|
脚註 및 參考 文獻
[
編輯
]