 酵素의 다른 자리 立體性(알로스테릭) 調節
酵素의 다른 자리 立體性(알로스테릭) 調節
다른 자리 立體性 調節
(
英語
:
allosteric regulation
)은
生化學
에서
酵素
의
活性 部位
가 아닌 다른 部位에
效果人者
와 結合하여 酵素를 調節하는 것이다.
알로스테릭 調節
,
알로스테릭 制御
(
英語
:
allosteric control
),
다른 자리 立體性 制御
라고도 한다.
效果因子가 結合하는 部位를 다른 자리 立體性 部位 또는 알로스테릭 部位(allosteric site) 또는 調節 部位(regulatory site)라고 한다. 다른 자리 立體性 部位는 效果因子가 蛋白質에 結合하도록 許容하여 種種
立體構造的 變化
및
蛋白質 動力學
의 變化를 招來한다.
[1]
[2]
蛋白質의 活性을 向上시키는 效果인자는 다른 자리 立體性 活性化劑(allosteric activator) 또는 알로스테릭 活性化劑라고 하며, 蛋白質의 活性을 감소시키는 效果인자는 다른 자리 立體性 沮害劑(allosteric inhibitor) 또는 알로스테릭 沮害劑라고 한다.
다른 자리 立體性 調節은 下流 生成物로부터의
피드백
또는 上流
氣質
의
피드포워드
와 같은 調節 루프의 자연스러운 例이다. 長距離 다른 자리 立體性 效果는
細胞 信號傳達
에 特히 重要하다.
[3]
다른 자리 立體性 調節은 또한
酵素
의 活性을 調節하는
細胞
의 能力에 있어서 特히 重要하다.
다른 자리 立體性 效果(allostery) 또는 알로스테릭 效果라는 用語는
古代 그리스어
에서 "其他(other)"를 意味하는 "allos (?λλο?)" 및 "固體(solid) 또는 對象(object)"을 意味하는 "stereos (στερε??)"에서 由來하였다. 이는 다른 자리 立體性 蛋白質의 調節 部位가 活性 部位와 物理的으로 다르다는 事實과 關聯이 있다. 알로스테릭 效果는 酵素의 活性化를 위해 立體構造的 變化가 必要하지 않은
氣質 提示
와 對照된다.
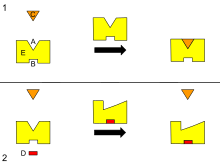 A ?
活性 部位
A ?
活性 部位
B ? 알로스테릭 部位
C ?
氣質
D ?
沮害劑
E ?
酵素
이것은 酵素의 알로스테릭 調節에 對한 模式圖이다.
많은 다른 자리 立體性 效果는
모노
,
와이만
,
찬듀
가 提示한
모노-와이만-찬듀 모델
(MWC 모델 또는 協同 모델이라고도 함)
[4]
또는
코쉬랜드
, 네메티(Nemethy), 필머(Filmer)가 說明한
順次 모델
(KNF 모델이라고도 함)로 說明할 수 있다.
[5]
둘 다
蛋白質 小段位體
가 緊張된(T) 또는 弛緩된(R)의 두 가지
立體構造
中 하나로 存在하며 弛緩된 小單位體가 緊張된 小段位體보다 氣質에 더 쉽게 結合한다고 假定한다. 이 두 가지 모델은 小段位體 相互作用과 두 狀態의 辭典 存在에 對한 家庭에서 가장 差異가 난다. 小單位體가 두 가지 以上의
立體構造
로 存在하는 蛋白質의 境遇 쿠엔데(Cuendet), 바인슈타인(Weinstein), 레빈(LeVine)李 說明한 알로스테리 地形 모델(allostery landscape model)을 使用할 수 있다.
[6]
다른 자리 立體性 調節은 大規模의 低에너지 立體構造的 變化의 進化에 依해 促進될 수 있으며, 이는 멀리 떨어진
結合 部位
사이의 長距離 다른 자리 立體性 相互作用을 可能하게 한다.
[7]
對稱 모델(symmetry model) 또는
MWC 모델
이라고도 하는 다른 자리 立體性 調節의 協同 모델(concerted model)은 어떤 소單位體의 立體構造的 變化가 必然的으로 다른 모든 소單位體에 附與되는 方式으로 酵素 小單位體가 連結되어 있다고 假定한다. 따라서 모든 소單位體는 同一한 立體構造로 存在해야 한다. 또한 이 모델은
리간드
(
氣質
또는 其他 分子)가 없는 境遇 平衡 狀態는 T 또는 R 立體構造적 狀態 中 하나를 選好한다고 主張한다. 하나의 리간드(다른 자리 立體性 效果人者 또는 리간드)가 活性 部位와 다른 部位에 結合함으로써 平衡이 R 또는 T 狀態로 轉換될 수 있다.
다른 자리 立體性 調節의 順次 모델(sequential model)은 어떤 소單位體의 立體構造的 變化가 다른 小段位體에서도 類似한 變化를 誘導하는 方式으로 小單位體가 連結되어 있지 않다고 主張한다. 따라서 모든 酵素 小單位體는 同一한 立體構造를 必要로 하지 않는다. 더욱이 順次 모델은 氣質 分子가
誘導 適合
프로토콜을 통해 結合함을 나타낸다. 이러한 誘導 適合은 小段位體를 緊張된 狀態에서 弛緩된 狀態로 變換시키지만 立體構造的 變化를 隣接한 小單位體로 傳播하지는 않는다. 代身 한 小段位體에서의 氣質 結合은 다른 小單位體의 構造를 若干만 變更하여 該當 小單位體의 結合 部位가 氣質을 더 잘 受容할 수 있도록 한다. 要約하면 다음과 같다.
- 小單位體는 同一한 立體構造로 存在할 必要가 없다.
- 氣質 分子는 誘導 適合 프로토콜을 통해 結合한다.
- 立體構造的 變化는 모든 소單位體에 傳播되지 않는다.
모르페인 모델
[
元本 編輯
]
다른 자리 立體性 調節의
모르페인
모델은 해리性 協同 모델이다.
[8]
모르페인은 生理學的으로 重要하고 機能的으로 다른 代替 4次 어셈블리의 앙상블로 存在할 수 있는 同種올리고머이다. 代替 모르페인 어셈블리 사이의 轉換에는 올리고머 海里, 해리 狀態의 立體構造的 變化 및 다른 올리고머로의 再組立이 包含된다. 必要한 올리고머 分解 段階는 다른 자리 立體性 調節을 위한 모르페인 모델을 旣存의 MWC 모델 및 KNF 모델과 差別化한다.
抛르포빌리노젠 生成酵素
(PBGS)는 모르페인의 프로토타입이다.
앙상블 모델
[
元本 編輯
]
다른 자리 立體性 調節의 앙상블 모델은 다른 자리 立體性 시스템의
統計的 앙상블
을 位置 에너지 函數의 函數로 列擧한 다음 다른 자리 立體性 效果의 特定 統計 測定값을 에너지 函數의 特定 에너지 要엉(예: 두 도메인 사이의 分子間 鹽다리)와 聯關시킨다.
[9]
앙상블 알로스테릭 모델(allosteric model)
[10]
및 알로스테릭 아이싱 모델(allosteric ising model)
[11]
과 같은 앙상블 모델은 시스템의 各 도메인이 WMC 모델과 類似한 두 가지 狀態를 採擇할 수 있다고 假定한다. 쿠엔데, 바인슈타인, 레빈이 導入한 알로스테릭 地形 모델
[6]
을 使用하면 도메인이 여러 狀態를 가질 수 있으며 주어진 알로스테릭 커플링에 對한 特定 分子 相互作用의 寄與度는 嚴格한 規則을 使用하여 推定할 수 있다.
分子動力學
시뮬레이션을 使用하면 시스템의 統計的 앙상블을 推定하여 알로스테리 地形 모델로 分析할 수 있다.
다른 자리 立體性 調節因子
[
元本 編輯
]
다른 자리 立體性 調節因子
(알로스테릭 調節因子)는 生化學 및 藥理學에서 分子와 酵素의 活性을 變更하는 데 使用된다. 比較를 위해 一般的인 藥물은 酵素의 活性 部位에 結合하여 氣質이 該當 酵素에 結合하는 것을 防止하여 酵素 活性을 감소시킨다. 알로스테릭 調節은
效果人者
가 酵素의 알로스테릭 部位(調節 部位라고도 函)에 結合하여 酵素의 活性을 變更할 때 일어난다. 알로스테릭 調節因子는 알로스테릭 部位에 맞게 設計되어 酵素의 立體構造的 變化, 特히 活性 部位의 形態 變化를 일으키고 이로 인해 酵素의 活性이 變化된다. 一般的인 藥물과 달리 調節因子는
競爭的 沮害劑
가 아니다. 이는 酵素 活性을 증가시키는 活性化(陽性) 또는 酵素 活性 減少를 誘發하는 沮害(陰性)일 수 있다. 알로스테릭 調節을 使用하면 特定 酵素의 活性의 效果를 制御할 수 있다. 結果的으로 알로스테릭 調節因子는 藥理學에서 매우 效果的이다.
[12]
生物學的 시스템에서 알로스테릭 調節은
氣質 提示
에 依한 調節과 區別하기 어려울 수 있다.
에너지 感知 모델
[
元本 編輯
]
에너지 感知 모델의 例는 사람의 大食細胞에 完璧하게 適應한
細菌
人
結核菌
(
Mycobacterium tuberculosis
)에서 볼 수 있다. 酵素의 部位는 서로 다른 氣質 間의 通信 役割을 한다. 特히,
AMP
와
葡萄糖 6-因山
(G6P) 사이에서 그러하다. 이와 같은 部位는 酵素 性能에 對한 感知 메커니즘 役割도 한다.
[13]
陽性 알로스테릭 調節(알로스테릭 活性化(allosteric activation)라고도 函)은 어떤
리간드
의 結合이 氣質 分子와 다른 結合 部位 사이의 人力을 强化시킬 때 일어난다. 例를 들어,
酸素
分子가
헤모글로빈
에 結合하는 境遇가 있는데, 여기서 酸素는 事實上
氣質
이자 效果因子이다. 알로스테릭 部位 또는 "其他" 部位는 隣接한
蛋白質 小段位體
의
活性 部位
이다. 하나의 小單位體에 酸素가 結合하면 該當 小單位體의 立體構造的 變化가 誘導되어 나머지 活性 部位와 相互作用하여 酸素 親和혁을 增加시킨다. 알로스테릭 活性化의 또 다른 例는 細胞質 IMP-GMP 特異的 5'-뉴클레오타이데이스 II(cN-II)에서 볼 수 있으며, 여기서 氣質인 GMP에 對한 親和力은
이合體
境界面에서 GTP 結合 時 增加한다.
陰性 알로스테릭 調節(알로스테릭 沮害(allosteric inhibition)라고도 函)은 어떤
리간드
의 結合이 다른 活性 部位의 氣質에 對한 親和力을 감소시킬 때 일어난다. 例를 들어,
2,3-비스포스포글리세르산
(2,3-BPG)李 헤모글로빈의 알로스테릭 部位에 結合하면 모든 소單位體의 酸素 親和力이 減少한다. 이는 結合 部位에 調節因子가 없는 境遇이다.
直接 트롬빈 沮害劑
는 陰性 알로스테릭 調節의 훌륭한 例이다. 潛在的으로
抗凝固劑
로 使用될 수 있는 트롬빈의 알로스테릭 沮害劑가 發見되었다.
또 다른 例는
글리신 受容體
의 알로스테릭 沮害劑 役割을 하는
痙攣性
독인
스트리크닌
이다.
글리신
은
哺乳類
의
脊髓
와
腦줄기
에서 主要
시냅스
後 抑制性
神經傳達物質
이다. 스트리크닌은 글리신 受容體의 別途 結合 部位에서 알로스테릭 方式으로 作用한다. 卽, 그 結合은 글리신에 對한 글리신 受容體의
親和性
을 낮춘다. 따라서 스트리크닌은 抑制性 傳達物質의 作用을 抑制하여 痙攣을 誘發한다.
陰性 알로스테릭 調節이 나타날 수 있는 또 다른 事例는
ATP
와
該當過程
을 調節하는
陰性 피드백
루프 內의 酵素인
포스포프럭토키네이스
사이에서 볼 수 있다. 포스포프럭토키네이스(일一般的로
PFK
라고 函)는 該當過程의 세 番째 段階인
果糖 6-因山
이
果糖 1,6-二重因山
으로
燐酸化
되는 過程을 觸媒하는 酵素이다.
포스포프럭토키네이스
는 細胞 내 높은 水準의
ATP
에 依해 알로스테릭하게 沮害될 수 있다. ATP의 水準이 높으면 ATP는 포스포프럭토키네이스의 알로스테릭 部位에 結合하여 酵素의 3次元的 形態의 變化를 일으킨다. 이러한 變化로 인해 活性 部位의
氣質
(
果糖 6-因山
및
ATP
)에 對한
親和力
이 減少하고 酵素가 非活性人 것으로 看做된다. 이는 ATP 水準이 높을 때
該當過程
을 中斷시켜 身體의
葡萄糖
을 保存하고 細胞의 ATP를 均衡잡힌 水準으로 維持한다. 이러한 方式으로 ATP는 酵素의 氣質임에도 不拘하고 포스포프럭토키네이스에 對한 音聲 알로스테릭 調節因子로 役割을 한다.
호모트로픽(homotropic) 알로스테릭 調節因子는 標的
蛋白質
의
氣質
이자 蛋白質 活性의 調節 分子이다. 이는 一般的으로 蛋白質의 活性化劑이다.
[14]
例를 들어, 酸素(O
2
)와 일酸化 炭素(CO)는 헤모글로빈의 알로스테릭 調節因子이다. 마찬가지로, IMP/GMP 特異的 5' 뉴클레오타이데이스에서 하나의 GMP 分子가 社合體 酵素의 單一 小單位體에 結合하면 S字形 氣質 代 速度 플롯에 依해 밝혀진 바와 같이 後續 小單位體에 依한 GMP에 對한 親和度를 증가시킨다.
[14]
헤테로트로픽
[
元本 編輯
]
헤테로트로픽(heterotropic) 알로스테릭 調節因子는 酵素의 氣質이 아닌 調節 分子이다. 이는 酵素의 活性化劑 또는 沮害劑日 수 있다. 例를 들어, H
+
, CO
2
및
2,3-비스포스포글리세르산
(2,3-BPG)은
헤모글로빈
의 헤테로트로픽 알로스테릭 調節因子이다.
[15]
IMP/GMP 特異的 5' 뉴클레오타이데이스에서 사량體 酵素의 異量體 境界面에서 GTP 分子의 結合은 K型 헤테로트로픽 알로스테릭 活性化를 나타내는 活性 部位에서 氣質인 GMP에 對한 親和力을 增加시킨다.
[14]
위에서 充分히 强調한 바와 같이, 一部 알로스테릭 蛋白質은 氣質과 다른 分子에 依해 調節될 수 있다. 이러한 蛋白質은 호모트로픽 相互作用과 헤테로트로픽 相互作用이 모두 可能하다.
[14]
必須 活性化劑
[
元本 編輯
]
一部 알로스테릭 活性化劑는 例를 들어, 카르바모일 因山 合成酵素 I에 對한
N
-아세틸글루탐산의 活性과 마찬가지로 不在時 標的 酵素의 活性이 매우 낮거나 無視할 수 있다는 意味에서 "必須的(essential)" 또는 "義務的(obligate)" 活性化劑라고 한다.
[16]
[17]
非調節 알로스테릭 效果
[
元本 編輯
]
非調節 알로스테릭 部位는 그 自體로는 아미노산이 아닌 酵素(또는 蛋白質)의 非調節 成分이다. 例를 들어, 많은 酵素들은 適切한 機能을 保障하기 위해 나트륨 結合을 必要로 한다. 그러나 나트륨이 반드시 調節 小單位體로 作用하는 것은 아니다. 나트륨은 恒常 存在하며 酵素 活性을 調節하기 위해 나트륨을 追加 또는 除去하는 것으로 알려진 生物學的 過程은 없다. 非調節 알로스테릭 調節은 나트륨 外에 다른 이온(칼슘, 마그네슘, 亞鉛)뿐만 아니라 다른 化合物과 비타민도 包含될 수 있다.
受容體의 알로스테릭 調節은
內因性 리간드
의 結合 部位(
活性 部位
)와 다른 部位(調節 部位)에서 알로스테릭 調節因子의 結合으로 인해 發生하며 內因性 리간드의 效果를 强化하거나 抑制한다. 正常的인 狀況에서는 受容體 分子의
立體構造的 變化
를 誘發하여 作用하며, 이로 因해 리간드의
結合 親和度
가 變更된다. 이러한 方式으로, 알로스테릭 리간드는 日次 오르토스테릭 리간드에 依해 受容體의 活性化를 調節하고 電氣 回路의 調光機 스위치처럼 作用하여 反應의 强度를 調整하는 것으로 생각할 수도 있다.
例를 들어,
GABA
A
受容體
에는 神經傳達物質인
γ-아미노뷰티르山
(GABA)李 結合하는 두 個의 活性 部位가 있을 뿐만 아니라
벤조다이아제핀
및
全身痲醉劑
調節 結合 部位도 있다. 이러한 調節 部位는 各各 養成 알로스테릭 調節을 하여 GABA의 活性을
强化
할 수 있다.
디아제팜
은 벤조디아제핀 調節 部位의
陽性 알로스테릭 調節因子
이며, 디아제팜의 解毒劑인
플壘禡祭닐
은
受容體 拮抗劑
이다.
標的을 알로스테릭하게 調節하는 藥物의 最近 例로는 不갑상샘機能亢進症 治療에 使用되는 藥물인
時나칼세트
와 HIV 感染 治療에 使用되는 藥물인
마라비록
이 있다.
藥物 標的으로서의 알로스테릭 部位
[
元本 編輯
]
알로스테릭 蛋白質은 많은 疾病에 關與하고 그 中心이 되며,
[18]
[19]
알로스테릭 部位는 새로운
藥物 標的
이 될 수 있다. 古典的인 오르토스테릭 리간드에 비해 選好되는 治療劑로서 알로스테릭 調節因子를 使用하는 데에는 많은 利點이 있다. 例를 들어,
G 蛋白質 連結 受容體
(GPCR)의 알로스테릭 結合 部位는 內因性 리간드를 受容하기 위해
오르토스테릭 部位
(
英語
:
orthosteric site
)와 같이 同一한 進化的 壓力에 直面하지 않았기 때문에 더 다양하다.
[20]
따라서 알로스테릭 部位를 標的으로 삼아 더 큰 G 蛋白質 連結 受容體(GPCR) 選擇性을 얻을 수 있다.
[20]
이는 受容體 下位 類型에 걸쳐 오르토스테릭 部位의 序列 保存으로 인해 選擇的 오르토스테릭 治療가 어려운 G 蛋白質 連結 受容體에 特히 有用하다.
[21]
또한 이러한 調節因子는 潛在的인 毒性 效果를 감소시킨다. 왜냐하면 協同性이 制限된 調節因子는 投與된 容量에 關係없이 效果가 上限線에 있기 때문이다.
[20]
알로스테릭 調節因子에 固有한 또 다른 類型의 藥理學的 選擇性은 協同性에 기초한다. 알로스테릭 調節因子는 關心있는 下位 類型을 除外하고 주어진 受容體의 모든 下位 類型에서 오르토스테릭 리간드와 中立的 協同性을 나타낼 수 있으며, 이를 "絶對 下位 類型 選擇性(absolute subtype selectivity)"이라고 한다.
[21]
알로스테릭 調節因子가 相當한 效能을 갖지 않는 境遇, 오르토스테릭 리간드에 비해 또 다른 强力한 治療 利點, 卽 內因性 作用제가 存在할 때만 組織 反應을 選擇的으로 調整하는 能力을 提供할 수 있다.
[21]
올리고머 特異的 低分子 結合 部位는 醫學的으로 關聯된
모르페인
의 藥物 標的이다.
[22]
合成 알로스테릭 시스템
[
元本 編輯
]
여러 個의
非共有 結合
部位를 包含하는 合成 化合物이 많이 있으며, 이는 한 部位를 차지할 때
立體構造的 變化
를 나타낸다. 그러한
超分子
시스템에서 單一 結合 寄與 사이의 協同性은 한 結合 部位의 占有가 두 番째 部位에서 親和力 Δ
G
를 높이는 境遇 兩性이고, 親和力이 높아지지 않으면 音聲이다. 大部分의 合成 알로스테릭 複合體는 하나의 效果人者 리간드의 結合에 따른 立體構造적 再構成에 依存하며, 이는 다른 結合 部位에서 두 番째 리간드의 結合을 强化하거나 약화시킨다.
[23]
[24]
[25]
여러 結合 部位 사이의 立體構造적 結合은 一般的으로 柔軟性이 더 큰 蛋白質보다 人工的인 시스템에서 훨씬 더 크다. 效率을 決定하는 媒介變數(效果人者 E의 存在 및 不在 下에서 平衡常數 Krel = KA(E)/KA의 比率로 測定됨)는 리간드 A의 結合을 위해 閉鎖된 또는 變形된 立體構造를 採擇하는 데 必要한 立體構造적 에너지이다.
[26]
많은 다가
超分子
시스템에서는
[27]
結合된 리간드 사이에 直接的인 相互作用이 일어날 수 있으며, 이는 큰 協同性을 誘發할 수 있다. 가장 一般的인 것은 이온쌍 受容體의 이온 間의 直接的인 相互作用이다.
[28]
[29]
이러한 協同性은 種種 알로스테릭 效果(allostery)라고도 불리지만, 여기서 立體構造的 變化가 반드시 結合 이벤트를 觸發하는 것은 아니다.
온라인 리소스
[
元本 編輯
]
알로스테릭 데이터베이스
[
元本 編輯
]
알로스테릭 調節은 오르토스테릭 部位와 地形的으로 區別되는 알로스테릭 部位에 리간드가 結合하여 生成되는 生物學的 巨大分子 機能의 調節을 위한 直接的이고 效率的인 手段이다. 種種 높은 受容體 選擇性과 낮은 標的 基盤 毒性으로 인해 알로스테릭 調節은 藥物 發見 및 生命工學에서 漸漸 더 많은 役割을 할 것으로 豫想된다.
알로스테릭 데이터베이스
(ASD)
[30]
는 알로스테릭 分子의 構造, 機能 및 關聯된 註釋을 標示, 檢索 및 分析하기 위한 中央 리소스를 提供한다. 現在 알로스테릭 데이터베이스에는 100種 以上의 알로스테릭 蛋白質과 세 가지 範疇의 調節因子(modulator) (活性化劑, 沮害劑, 調節因子(regulator))가 包含되어 있다. 各 蛋白質에는 알로스테리(allostery), 生物學的 過程 및 關聯 疾病에 對한 仔細한 說明과 結合 親和度, 物理化學的 特性 및 治療 領域이 있는 各 調節因子에 對한 註釋이 달려 있다. 알로스테릭 데이터베이스에 알로스테릭 蛋白質의 情報를 統合하면 알려지지 않은 蛋白質에 對한 알로스테리(알로스테릭 效果)를 豫測하고 實驗的 檢證을 遂行할 수 있다. 또한 알로스테릭 데이터베이스에서 選別된 調節因子는 問議된 化合物에 對한 潛在的인 알로스테릭 標的을 調査하는 데 使用될 수 있으며 化學者가 새로운 알로스테릭 藥物 設計를 위한 構造 變形을 具現하는 데 도움이 될 수 있다.
알로스테릭 殘基 및 그 豫測
[
元本 編輯
]
모든 蛋白質 殘氣가 알로스테릭 調節에서 똑같이 重要한 役割을 하는 것은 아니다. 알로스테리에 必須的인 殘基(所謂 "알로스테릭 殘基")의 識別은 特히 지난 10年 동안 많은 硏究의 焦點이 되어 왔다.
[31]
[32]
[33]
[34]
[35]
[36]
[37]
[38]
部分的으로 이러한 關心의 增加는 蛋白質 科學에서의 一般的인 重要性 때문일 뿐만 아니라 알로스테릭 殘氣가 生醫學的 脈絡에서 活用될 수 있기 때문이다. 標的化하기 어려운 部位를 가진 藥理學的으로 重要한 蛋白質은 關心 있는 1次 部位를 알로스테릭하게 調節할 수 있는 到達하기 쉬운 殘氣를 對案的으로 標的化하는 接近 方式을 提供할 수 있다.
[39]
이러한 殘期는 廣範圍하게 表面 알로스테릭 아미노산 및 內部 알로스테릭 아미노산으로 分類될 수 있다. 表面 알로스테릭 部位는 一般的으로 內部 알로스테릭 部位와 根本的으로 區別되는 調節 役割을 한다. 表面 殘期는 알로스테릭 信號傳達에서 受容體 또는 效果記 部位 役割을 할 수 있는 反面, 內部 殘期는 그러한 信號를 傳達하는 役割을 할 수 있다.
[40]
[41]
- ↑
Cooper A, Dryden DT (October 1984). “Allostery without conformational change. A plausible model”. 《European Biophysics Journal》
11
(2): 103?109.
doi
:
10.1007/BF00276625
.
PMID
6544679
.
S2CID
12591175
.
- ↑
Liu J, Nussinov R (June 2016).
“Allostery: An Overview of Its History, Concepts, Methods, and Applications”
. 《PLOS Computational Biology》
12
(6): e1004966.
Bibcode
:
2016PLSCB..12E4966L
.
doi
:
10.1371/journal.pcbi.1004966
.
PMC
4890769
.
PMID
27253437
.
S2CID
3610740
.
- ↑
Bu Z, Callaway DJ (2011). 〈Proteins move! Protein dynamics and long-range allostery in cell signaling〉. 《Protein Structure and Diseases》. Advances in Protein Chemistry and Structural Biology
83
. 163?221쪽.
doi
:
10.1016/B978-0-12-381262-9.00005-7
.
ISBN
9780123812629
.
PMID
21570668
.
- ↑
Monod J, Wyman J, Changeux JP (May 1965). “On the nature of allosteric transitions:A plausible model”. 《Journal of Molecular Biology》
12
: 88?118.
doi
:
10.1016/s0022-2836(65)80285-6
.
PMID
14343300
.
- ↑
Koshland DE, Nemethy G, Filmer D (January 1966). “Comparison of experimental binding data and theoretical models in proteins containing subunits”. 《Biochemistry》
5
(1): 365?85.
doi
:
10.1021/bi00865a047
.
PMID
5938952
.
- ↑
가
나
Cuendet MA, Weinstein H, LeVine MV (December 2016).
“The Allostery Landscape: Quantifying Thermodynamic Couplings in Biomolecular Systems”
. 《Journal of Chemical Theory and Computation》
12
(12): 5758?5767.
doi
:
10.1021/acs.jctc.6b00841
.
PMC
5156960
.
PMID
27766843
.
- ↑
Eckmann JP, Rougemont J, Tlusty T (2019年 7月 30日).
“Colloquium : Proteins: The physics of amorphous evolving matter”
. 《Reviews of Modern Physics》 (英語)
91
(3): 031001.
arXiv
:
1907.13371
.
Bibcode
:
2019RvMP...91c1001E
.
doi
:
10.1103/RevModPhys.91.031001
.
ISSN
0034-6861
.
S2CID
199001124
.
- ↑
Jaffe EK (September 2005). “Morpheeins--a new structural paradigm for allosteric regulation”. 《Trends in Biochemical Sciences》
30
(9): 490?7.
doi
:
10.1016/j.tibs.2005.07.003
.
PMID
16023348
.
- ↑
Motlagh HN, Wrabl JO, Li J, Hilser VJ (April 2014).
“The ensemble nature of allostery”
. 《Nature》
508
(7496): 331?9.
Bibcode
:
2014Natur.508..331M
.
doi
:
10.1038/nature13001
.
PMC
4224315
.
PMID
24740064
.
- ↑
Hilser VJ, Wrabl JO, Motlagh HN (2012).
“Structural and energetic basis of allostery”
. 《Annual Review of Biophysics》
41
: 585?609.
doi
:
10.1146/annurev-biophys-050511-102319
.
PMC
3935618
.
PMID
22577828
.
- ↑
LeVine MV, Weinstein H (May 2015).
“AIM for Allostery: Using the Ising Model to Understand Information Processing and Transmission in Allosteric Biomolecular Systems”
. 《Entropy》
17
(5): 2895?2918.
Bibcode
:
2015Entrp..17.2895L
.
doi
:
10.3390/e17052895
.
PMC
4652859
.
PMID
26594108
.
- ↑
Abdel-Magid AF (February 2015).
“Allosteric modulators: an emerging concept in drug discovery”
. 《ACS Medicinal Chemistry Letters》
6
(2): 104?7.
doi
:
10.1021/ml5005365
.
PMC
4329591
.
PMID
25699154
.
- ↑
Zhong W, Cui L, Goh BC, Cai Q, Ho P, Chionh YH, Yuan M, Sahili AE, Fothergill-Gilmore LA, Walkinshaw MD, Lescar J, Dedon PC (December 2017).
“Allosteric pyruvate kinase-based "logic gate" synergistically senses energy and sugar levels in Mycobacterium tuberculosis”
. 《Nature Communications》
8
(1): 1986.
Bibcode
:
2017NatCo...8.1986Z
.
doi
:
10.1038/s41467-017-02086-y
.
PMC
5719368
.
PMID
29215013
.
- ↑
가
나
다
라
Srinivasan B, Forouhar F, Shukla A, Sampangi C, Kulkarni S, Abashidze M, Seetharaman J, Lew S, Mao L, Acton TB, Xiao R, Everett JK, Montelione GT, Tong L, Balaram H (March 2014).
“Allosteric regulation and substrate activation in cytosolic nucleotidase II from Legionella pneumophila”
. 《The FEBS Journal》
281
(6): 1613?1628.
doi
:
10.1111/febs.12727
.
PMC
3982195
.
PMID
24456211
.
- ↑
Edelstein SJ (1975). “Cooperative interactions of hemoglobin”. 《Annual Review of Biochemistry》
44
: 209?32.
doi
:
10.1146/annurev.bi.44.070175.001233
.
PMID
237460
.
- ↑
Shi D, Allewell NM, Tuchman M (June 2015).
“The N-Acetylglutamate Synthase Family: Structures, Function and Mechanisms”
. 《International Journal of Molecular Sciences》
16
(6): 13004?22.
doi
:
10.3390/ijms160613004
.
PMC
4490483
.
PMID
26068232
.
- ↑
de Cima S, Polo LM, Diez-Fernandez C, Martinez AI, Cervera J, Fita I, Rubio V (November 2015).
“Structure of human carbamoyl phosphate synthetase: deciphering the on/off switch of human ureagenesis”
. 《Scientific Reports》
5
(1): 16950.
Bibcode
:
2015NatSR...516950D
.
doi
:
10.1038/srep16950
.
PMC
4655335
.
PMID
26592762
.
- ↑
Nussinov R, Tsai C (2013). “Allostery in disease and in drug discovery”. 《Cell》
153
(2): 293?305.
doi
:
10.1016/j.cell.2013.03.034
.
PMID
23582321
.
- ↑
Abrusan G, Ascher DB, Inouye M (2022). “Known allosteric proteins have central roles in genetic disease”. 《PLOS Computational Biology》
18
(2): e1009806.
arXiv
:
2107.04318
.
Bibcode
:
2022PLSCB..18E9806A
.
doi
:
10.1371/journal.pcbi.1009806
.
PMID
10138267
.
- ↑
가
나
다
Christopoulos A, May LT, Avlani VA, Sexton PM (November 2004). “G-protein-coupled receptor allosterism: the promise and the problem(s)”. 《Biochemical Society Transactions》
32
(Pt 5): 873?7.
doi
:
10.1042/BST0320873
.
PMID
15494038
.
- ↑
가
나
다
May LT, Leach K, Sexton PM, Christopoulos A (2007). “Allosteric modulation of G protein-coupled receptors”. 《Annual Review of Pharmacology and Toxicology》
47
: 1?51.
doi
:
10.1146/annurev.pharmtox.47.120505.105159
.
PMID
17009927
.
- ↑
Jaffe EK (2010).
“Morpheeins ? A New Pathway for Allosteric Drug Discovery~!2010-02-12~!2010-05-21~!2010-06-08~!”
. 《The Open Conference Proceedings Journal》
1
: 1?6.
doi
:
10.2174/2210289201001010001
(年 以後로 接續 不可 2024-03-11).
PMC
3107518
.
PMID
21643557
.
- ↑
Takeuchi M, Ikeda M, Sugasaki A, Shinkai S (November 2001). “Molecular design of artificial molecular and ion recognition systems with allosteric guest responses”. 《Accounts of Chemical Research》
34
(11): 865?73.
doi
:
10.1021/ar0000410
.
PMID
11714258
.
- ↑
Kremer C, Lutzen A (May 2013). “Artificial allosteric receptors”. 《Chemistry: A European Journal》
19
(20): 6162?96.
doi
:
10.1002/chem.201203814
.
PMID
23463705
.
- ↑
Kovbasyuk L, Kramer R (June 2004). “Allosteric supramolecular receptors and catalysts”. 《Chemical Reviews》
104
(6): 3161?87.
doi
:
10.1021/cr030673a
.
PMID
15186190
.
- ↑
Schneider HJ (September 2016). “Efficiency parameters in artificial allosteric systems”. 《Organic & Biomolecular Chemistry》
14
(34): 7994?8001.
doi
:
10.1039/c6ob01303a
.
PMID
27431438
.
- ↑
Badji? JD, Nelson A, Cantrill SJ, Turnbull WB, Stoddart JF (September 2005). “Multivalency and cooperativity in supramolecular chemistry”. 《Accounts of Chemical Research》
38
(9): 723?32.
doi
:
10.1021/ar040223k
.
PMID
16171315
.
- ↑
Kim SK, Sessler JL (October 2010).
“Ion pair receptors”
. 《Chemical Society Reviews》
39
(10): 3784?809.
doi
:
10.1039/c002694h
.
PMC
3016456
.
PMID
20737073
.
- ↑
McConnell AJ, Beer PD (May 2012). “Heteroditopic receptors for ion-pair recognition”. 《Angewandte Chemie》
51
(21): 5052?61.
doi
:
10.1002/anie.201107244
.
PMID
22419667
.
- ↑
Huang Z, Zhu L, Cao Y, Wu G, Liu X, Chen Y, Wang Q, Shi T, Zhao Y, Wang Y, Li W, Li Y, Chen H, Chen G, Zhang J (January 2011).
“ASD: a comprehensive database of allosteric proteins and modulators”
. 《Nucleic Acids Research》
39
(Database issue): D663?9.
doi
:
10.1093/nar/gkq1022
.
PMC
3013650
.
PMID
21051350
.
- ↑
Panjkovich A, Daura X (October 2012).
“Exploiting protein flexibility to predict the location of allosteric sites”
. 《BMC Bioinformatics》
13
: 273.
doi
:
10.1186/1471-2105-13-273
.
PMC
3562710
.
PMID
23095452
.
- ↑
Suel GM, Lockless SW, Wall MA, Ranganathan R (January 2003). “Evolutionarily conserved networks of residues mediate allosteric communication in proteins”. 《Nature Structural Biology》
10
(1): 59?69.
doi
:
10.1038/nsb881
.
PMID
12483203
.
S2CID
67749580
.
- ↑
Mitternacht S, Berezovsky IN (September 2011).
“Binding leverage as a molecular basis for allosteric regulation”
. 《PLOS Computational Biology》
7
(9): e1002148.
Bibcode
:
2011PLSCB...7E2148M
.
doi
:
10.1371/journal.pcbi.1002148
.
PMC
3174156
.
PMID
21935347
.
- ↑
Gasper PM, Fuglestad B, Komives EA, Markwick PR, McCammon JA (December 2012).
“Allosteric networks in thrombin distinguish procoagulant vs. anticoagulant activities”
. 《Proceedings of the National Academy of Sciences of the United States of America》
109
(52): 21216?22.
doi
:
10.1073/pnas.1218414109
.
PMC
3535651
.
PMID
23197839
.
- ↑
Ghosh A, Vishveshwara S (November 2008). “Variations in clique and community patterns in protein structures during allosteric communication: investigation of dynamically equilibrated structures of methionyl tRNA synthetase complexes”. 《Biochemistry》
47
(44): 11398?407.
doi
:
10.1021/bi8007559
.
PMID
18842003
.
- ↑
Sethi A, Eargle J, Black AA, Luthey-Schulten Z (April 2009).
“Dynamical networks in tRNA:protein complexes”
. 《Proceedings of the National Academy of Sciences of the United States of America》
106
(16): 6620?5.
Bibcode
:
2009PNAS..106.6620S
.
doi
:
10.1073/pnas.0810961106
.
PMC
2672494
.
PMID
19351898
.
- ↑
Vanwart AT, Eargle J, Luthey-Schulten Z, Amaro RE (August 2012).
“Exploring residue component contributions to dynamical network models of allostery”
. 《Journal of Chemical Theory and Computation》
8
(8): 2949?2961.
doi
:
10.1021/ct300377a
.
PMC
3489502
.
PMID
23139645
.
- ↑
Rivalta I, Sultan MM, Lee NS, Manley GA, Loria JP, Batista VS (May 2012).
“Allosteric pathways in imidazole glycerol phosphate synthase”
. 《Proceedings of the National Academy of Sciences of the United States of America》
109
(22): E1428?36.
doi
:
10.1073/pnas.1120536109
.
PMC
3365145
.
PMID
22586084
.
- ↑
Negre CF, Morzan UN, Hendrickson HP, Pal R, Lisi GP, Loria JP, Rivalta I, Ho J, Batista VS (December 2018).
“Eigenvector centrality for characterization of protein allosteric pathways”
. 《Proceedings of the National Academy of Sciences of the United States of America》
115
(52): E12201?E12208.
arXiv
:
1706.02327
.
Bibcode
:
2018PNAS..11512201N
.
doi
:
10.1073/pnas.1810452115
.
PMC
6310864
.
PMID
30530700
.
- ↑
Clarke D, Sethi A, Li S, Kumar S, Chang RW, Chen J, Gerstein M (May 2016).
“Identifying Allosteric Hotspots with Dynamics: Application to Inter- and Intra-species Conservation”
. 《Structure》
24
(5): 826?837.
doi
:
10.1016/j.str.2016.03.008
.
PMC
4883016
.
PMID
27066750
.
- ↑
Dutta S, Eckmann JP, Libchaber A, Tlusty T (May 2018).
“Green function of correlated genes in a minimal mechanical model of protein evolution”
. 《Proceedings of the National Academy of Sciences of the United States of America》
115
(20): E4559?E4568.
arXiv
:
1801.03681
.
Bibcode
:
2018PNAS..115E4559D
.
doi
:
10.1073/pnas.1716215115
.
PMC
5960285
.
PMID
29712824
.