C4 植物
은 4炭糖(C4) 化合物이 關與하는 追加的인 經路(C4回로)를 利用해
 가 不足한 環境에서도
光合成
의 暗反應(광비依存性反應)을 繼續할 수 있는 植物을 말한다. 4炭糖人 옥살아세트산이 最初의 炭素固定産物이기 때문에 "C4"經路 라는 이름이 붙여졌으며 1966年에 2名의 濠洲 科學者들 (Hatch& Slack)李 밝혀내었기 때문에 Hatch-Slack 經路라고도 불린다.
캘빈回路
(C3回로)의 Rubisco보다
를 效率的으로 固定하는 C4 植物의 炭素固定回路는
의 最初 固定 産物이 4炭糖(C4)인 데서 이름이 由來되었다. 大部分의
熱帶
或은
亞熱帶
性 植物이 이 무리에 屬한다.
가 不足한 環境에서도
光合成
의 暗反應(광비依存性反應)을 繼續할 수 있는 植物을 말한다. 4炭糖人 옥살아세트산이 最初의 炭素固定産物이기 때문에 "C4"經路 라는 이름이 붙여졌으며 1966年에 2名의 濠洲 科學者들 (Hatch& Slack)李 밝혀내었기 때문에 Hatch-Slack 經路라고도 불린다.
캘빈回路
(C3回로)의 Rubisco보다
를 效率的으로 固定하는 C4 植物의 炭素固定回路는
의 最初 固定 産物이 4炭糖(C4)인 데서 이름이 由來되었다. 大部分의
熱帶
或은
亞熱帶
性 植物이 이 무리에 屬한다.
는
空氣
中에 아주 적은 比率(0.03%)로 存在하며 追加的으로 溫度가 높고 日照量이 많은 環境의 條件에서
使用이 不安定한 狀態가 되면 캘빈回路에 있는
固定酵素인 Rubisco는 더 以上 炭素 固定보다는
光呼吸
(photorespiration)을 하는 比率이 높아지게 된다. 反面에 C4植物은 Rubisco보다 强力한
固定酵素인
PEPC
(Phosphoenolpyruvate carboxylase)를 利用해 炭素 固定을 繼續할 수 있다.
C3 植物과 C4 植物의 差異
[
編輯
]
C3 植物에 比해 C4 植物은 더 乾燥하고 溫度가 높으며 窒素源이 不足한 곳에서도 光合成을 할 수 있다. 이는 C4街 C3와는 다르기 때문에 可能한 것이다. 構造的인 差異와 臺詞 過程의 差異로 나누어 살펴보도록 하겠다.
構造的 差異
[
編輯
]
維管束(체管, 물管)은 2個의 세포층으로 둘러싸인 特徵的인 構造를 가지고 있다. 안쪽 細胞層은 有關束草(稍)細胞(bundle sheath cell;잎脈을 에워싸고 있는 細胞)라고 불리며 그라나 構造가 不足한 代身 澱粉이 많이 貯藏되어 있다. 바깥쪽 細胞層은 葉肉細胞(mesophyll cell)라고 불리며 維管束超細胞와는 反對로 澱粉 貯藏을 많이 하지 않지만 그라나 構造가 發達되어 있다. 안쪽과 바깥쪽의 같은
葉綠體
가 다른 形態를 나타낸다.
[1]
C4 植物 잎의 橫斷面을 觀察하면,
維管束超細胞
(bundle sheath cell)가 維管束의 周圍를 둘러싸고 있으며 그 周圍를 葉肉 細胞가 둘러싸고 있다. 이것은 마치 꽃다발처럼 보이므로, 크란츠 救助(Kranz anatomy)라고 한다.(Kranz
[2]
-Crown/Halo)라고 한다. C3 植物에서는 葉綠體가 葉肉 細胞에는 많지만, 有關束草 細胞에서는 적고 維管束草家 C4 植物만큼 두텁게 發達되어 있지 않아서 이러한 크란츠 構造가 나타나지 않는다. 그러나, C4 植物에서는 有關束草 細胞에도 葉綠體가 많이 存在하며 維管束秒가 두텁게 發達되어 크란츠 構造가 뚜렷하게 나타나는 것이 特徵이다.
C3 植物
에서는
葉肉細胞
(葉肉細胞, leaf mesopyll cell)에서 直接 CO2
와 接觸이 일어나 光合成을 하지만, C4 植物의 葉肉細胞에서는
를 有機酸의 形態로 固定하여
維管束
超細胞로 들여보내기 때문에 間接的으로
와 接觸하여 光合成을 進行하게 된다.
 의 境遇도 마찬가지로 C3 植物에서는 光合成이 일어나는 곳과 直接 接觸이 일어나지만 C4 植物에서는 間接的으로 接觸이 일어나기 때문에 光呼吸(높은 酸素 濃度 때문에 生成된 有機物이 分解되는 植物에게 害로운 過程)의 機會는 줄어들고 잎 內部의
濃度를 높여 炭素固定量을 늘릴 수 있다.
의 境遇도 마찬가지로 C3 植物에서는 光合成이 일어나는 곳과 直接 接觸이 일어나지만 C4 植物에서는 間接的으로 接觸이 일어나기 때문에 光呼吸(높은 酸素 濃度 때문에 生成된 有機物이 分解되는 植物에게 害로운 過程)의 機會는 줄어들고 잎 內部의
濃度를 높여 炭素固定量을 늘릴 수 있다.
代謝 過程
[
編輯
]
C3 植物
에서는
葉肉細胞
(leaf mesopyll cell)에서 캘빈回路가 돌아가지만, C4 植物에서는
維管束
超細胞(bundle sheath cell; 잎脈을 에워싸고 있는 細胞)에서 캘빈回路가 作動하고 葉肉細胞에서는 C4回路가 進行된다. PEP carboxylase에 依해 CO
2
가 固定되어 4炭素化合物이 되고, 이 回路의 最終産物인 말산(malate)은 葉肉細胞에서 隣接한 維管束超細胞로 移動하여 3炭素 化合物과 CO
2
로 分解된다. CO
2
는 Rubisco와 結合하여 維管束超細胞의 캘빈回路에 들어간다. 卽 相對的으로 性能이 나쁜 Rubisco 代身에 좀 더 强力한 酵素인 PEP carboxylase가 CO
2
를 代身 받아 줌으로써 낮은 CO
2
環境에서도 暗反應을 繼續 進行시킬 수 있는 것이다.
C4 植物은 一般的인 植物인 C3 植物에 比해, 發達된 有關束草 細胞가 있으며 葉綠體가 多數 存在한다. 이 때문에 C3 植物의 캘빈-벤슨 回路는 葉肉 細胞에 있는 데 비하여 C4 植物은 有關束草 細胞에 있다. C3 植物은 CO
2
를 固定하는 데 RubisCO를 利用하지만 C4 植物은 PEPC를 利用한다. C4 植物의 이러한 差異는 光呼吸을 줄여줘 光合成에 有利하게 作用한다. 普通 C3 植物의 CO
2
補償點은 40~100 ppm이지만, 高溫이 되면 上昇하여 補償點이 大氣 中의 CO
2
濃度(350 ppm)에 가까워진다. 이 때문에 高溫에서 C3 植物은 成長 速度가 制限될 可能性이 높아진다. 反面, C4 植物의 CO
2
補償點은 2~5 ppm으로 溫度가 높아져도 補償點이 C3만큼 急激히 增加하지는 않는다.
C4 植物은 C3 植物에 비해 半乾燥 條件에서도 잘 사는데 數分 使用率(光合成에 利用하는 물/增産으로 잃는 물)李 높기 때문이다.
C4 植物은 C3 植物에 비하여 窒素 利用 效率도 높다.
첫째, RubisCO의 生成量이 적은 것을 理由로 들 수 있다. C4 經路에 依한 CO
2
濃縮 期作에서 RubisCO의 酸化酵素(oxygenase)反應(光呼吸)李 거의 일어나지 않게 되기 때문에 RubisCO의 生成量이 적어도 된다. C3 植物에서 全體 蛋白質의 約 50%를 차지하는 RubisCO의 生成量을 줄일 수 있으므로 C4 植物은 窒素 利用 效率이 높아지게 된다.
둘째, 光呼吸에 依한 窒素의 再放出이 일어나지 않는다. C4 植物은 C3 植物에 比해, 광利用 效率이 높다. 强한 빛은
狂界
II, I(photo system II,I)의 不必要한 電子여기를 促進하여
活性酸素
를 發生시키므로 植物에 毒性을 나타내기 때문에 植物은 强한 빛에 對한 防禦를 한다. 빛에너지를 螢光(빛)이나 熱, 光呼吸을 통해 放出하거나 여러 가지 補助 色素(
카로티노이드
, 크립토크롬, 플라보노이드 等)로 强한 빛을 吸收하여 自己自身을 保護한다.
以上에서 본 바와 같이 高溫, 乾燥, 强한 빛, 窒素 不足 土壤 條件에서는 C4 植物이 C3 植物에 비해 유리하다. 또한 C4 植物은 最適條件日 때 C3 植物에 비하여 效率的으로 炭酸 固定이 進行되기 때문에, 같은 量의 빛에서 더 많은 炭素를 固定할 수 있다. 그러나 C4 經路에서 CO
2
固定에 ATP 2 分子가 더 消耗되므로 光呼吸이 적은 低溫의 濕潤한 地域에서는 C3 植物이 유리하기 때문에 C3 植物이 全 世界 植物의 大部分을 이루고 있다.
C4 植物의 分類
[
編輯
]
葉肉 細胞에서 PEP(3炭糖)에서
(實在 HCO3-)를 固精해 옥살산(4炭糖)을 만들어 다양한 物質로 變換되어 維管束束草 細胞에서 脫炭酸 酵素에 依해
가 재방출되어 캘빈-벤슨 回路에 받아들여지게 된다. C4 經路는 有關束草 細胞로
를 再放出할 때 關與하는 酵素(脫炭酸酵素)에 따라 크게 3 種類로 나뉜다.
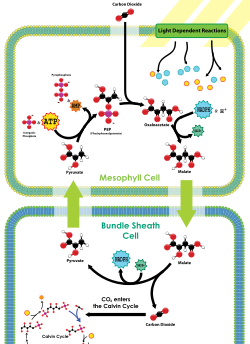 C4(Hatch-Slack) 經路- NADP-ME型
C4(Hatch-Slack) 經路- NADP-ME型
NADP-ME 兄
[
編輯
]
옥수수
,
沙糖수수
, 참억새 等이 包含된다. NADP-말산酵素(NADP-ME: NADP-malic enzyme)가 脫炭酸作用(decarboxylation)을 한다. 葉肉 細胞 氣質에서
가 물에 녹아
 가 되고 PEP와 PEP carboxylase(PEPC)에 依해 옥살아세트산이 된다. 옥살아세트산은 葉綠體에 들어가 말산脫水素酵素(MDH: malate dehydrogenase)와 還元劑 NADPH에 依해 4炭糖人 말산(沙果酸)으로 還元된다. 말産銀 葉綠體에서 細胞質로 放出되어 濃度 勾配에 依해 原形質 連絡士를 經由해, 有關束草 細胞로 移動한다. 有關束草 細胞의 葉綠體로 말산이 移動하여 NADP+와 脫炭酸酵素와 反應해 피루브산(3炭糖), NADPH,
이 生成된다. 脫炭酸過程에 關與하는 酵素가 NADP-말산酵素(NADP-ME: NADP-malic enzyme)이다. 여기서 生成된
는
캘빈-벤슨 回로
에 들어가며, 피루브산은 有關束草 細胞의 細胞質로 放出되어 濃度 勾配에 따라 原形質 連絡士를 거쳐 葉肉細胞의 葉綠體로 移動한다. 이 피루브산은, 2ATP를 利用해 다시 PEP가 되는데 이 反應에 關與하는 酵素를 피루브산因山 디키나아제(PPDK: pyruvate-phosphate dikinase)라고 한다. PPDK는 피루브산 1分子와 武器 燐酸 1分子, ATP 1分子의 에너지를 使用해 PEP 1分子와 AMP 1分子 無機燐酸 2分子를 形成한다. 이 AMP를 ADP로 되돌리기 위해 ATP 1分子가 使用된다. 따라서 NADP-ME形에서는 1 分子의
濃縮에 2 分子의 ATP가 必要하다.
가 되고 PEP와 PEP carboxylase(PEPC)에 依해 옥살아세트산이 된다. 옥살아세트산은 葉綠體에 들어가 말산脫水素酵素(MDH: malate dehydrogenase)와 還元劑 NADPH에 依해 4炭糖人 말산(沙果酸)으로 還元된다. 말産銀 葉綠體에서 細胞質로 放出되어 濃度 勾配에 依해 原形質 連絡士를 經由해, 有關束草 細胞로 移動한다. 有關束草 細胞의 葉綠體로 말산이 移動하여 NADP+와 脫炭酸酵素와 反應해 피루브산(3炭糖), NADPH,
이 生成된다. 脫炭酸過程에 關與하는 酵素가 NADP-말산酵素(NADP-ME: NADP-malic enzyme)이다. 여기서 生成된
는
캘빈-벤슨 回로
에 들어가며, 피루브산은 有關束草 細胞의 細胞質로 放出되어 濃度 勾配에 따라 原形質 連絡士를 거쳐 葉肉細胞의 葉綠體로 移動한다. 이 피루브산은, 2ATP를 利用해 다시 PEP가 되는데 이 反應에 關與하는 酵素를 피루브산因山 디키나아제(PPDK: pyruvate-phosphate dikinase)라고 한다. PPDK는 피루브산 1分子와 武器 燐酸 1分子, ATP 1分子의 에너지를 使用해 PEP 1分子와 AMP 1分子 無機燐酸 2分子를 形成한다. 이 AMP를 ADP로 되돌리기 위해 ATP 1分子가 使用된다. 따라서 NADP-ME形에서는 1 分子의
濃縮에 2 分子의 ATP가 必要하다.
NAD-ME 兄
[
編輯
]
機張
, 조 等이 包含된다. NAD-ME(NAD-말산酵素)가 脫炭酸作用을 한다. 于先 葉肉 細胞에서 받아들인
 는
는
 이온이 되며 PEP와 反應하여 옥살아세트산이 形成된다. 이때 PEPC가 觸媒로 作用한다. 獄事 아세트산에 아민基가 붙어 아스파르트산이 되면, 原形質 連絡士를 통해 濃度 差異에 따라 有關束草 細胞로 移動하여 미토콘드리아로 들어간다. 아스파르트산은 脫아민反應에 依해 다시 옥살아세트산이 된다. 옥살아세트산은 NADH에 依해 말산으로 還元되며 말産銀 NAD-ME(NAD-말산酵素)에 依해 脫炭酸 反應이 일어나 NADH, 피루브산,
를 生成한다.
는 擴散에 依해 葉綠體로 移動하여 캘빈-벤슨 回路로 들어가 다시 固定된다. 피루브산은 미토콘드리아에서 細胞質 氣質로 放出되면 아민基가 붙어 알라닌이 된다. 알라닌은 濃度 差異에 따라 原形質 連絡士를 거쳐 葉肉 細胞로 移動한다. 葉肉 細胞에서 알라닌은 다시 脫아민化되어 피루브산形態로 葉綠體에 들어간다. 그 以後는 NADP-ME型과 同一하게 進行되며 2 分子의 ATP 에너지를 使用해 PEP로 돌아온다. NAD-ME型 또한 1 分子의
農畜에는 2 分子의 ATP가 必要하다.
이온이 되며 PEP와 反應하여 옥살아세트산이 形成된다. 이때 PEPC가 觸媒로 作用한다. 獄事 아세트산에 아민基가 붙어 아스파르트산이 되면, 原形質 連絡士를 통해 濃度 差異에 따라 有關束草 細胞로 移動하여 미토콘드리아로 들어간다. 아스파르트산은 脫아민反應에 依해 다시 옥살아세트산이 된다. 옥살아세트산은 NADH에 依해 말산으로 還元되며 말産銀 NAD-ME(NAD-말산酵素)에 依해 脫炭酸 反應이 일어나 NADH, 피루브산,
를 生成한다.
는 擴散에 依해 葉綠體로 移動하여 캘빈-벤슨 回路로 들어가 다시 固定된다. 피루브산은 미토콘드리아에서 細胞質 氣質로 放出되면 아민基가 붙어 알라닌이 된다. 알라닌은 濃度 差異에 따라 原形質 連絡士를 거쳐 葉肉 細胞로 移動한다. 葉肉 細胞에서 알라닌은 다시 脫아민化되어 피루브산形態로 葉綠體에 들어간다. 그 以後는 NADP-ME型과 同一하게 進行되며 2 分子의 ATP 에너지를 使用해 PEP로 돌아온다. NAD-ME型 또한 1 分子의
農畜에는 2 分子의 ATP가 必要하다.
PEP-CK 兄
[
編輯
]
PCK兄이라고도 한다. PEP-CK型은 脫炭酸 酵素에 PEP-CK(PEP-carboxykinase)가 使用된다. NAD-ME과 類似한 듯 보이나 有關束草 細胞의 미토콘드리아가 아닌 葉綠體에서 反應이 일어나는 것과 中間産物 一部가 다르다. 먼저 葉肉 細胞에서 받아들인
는
이온이 되며 PEP와 反應하여 옥살아세트산이 形成된다. 이때 PEPC가 觸媒로 作用한다. 獄事 아세트산에 아민基가 붙어 아스파르트산이 되면, 原形質 連絡士를 통해 濃度 差異에 따라 有關束草 細胞로 移動하여 葉綠體로 들어간다. 아스파르트산은 脫아민反應에 依해 다시 옥살아세트산이 된다. 옥살아세트산은 ATP에 依해 PEP와
로 쪼개어지며
는 擴散에 依해 葉綠體로 移動하여 캘빈-벤슨 回路로 들어간다. PEP는 細胞質 氣質로 放出되면 아민基가 붙어 알라닌이 된다. 알라닌은 濃度 差異에 따라 原形質 連絡士를 거쳐 葉肉 細胞로 移動한다. 葉肉 細胞에서 알라닌은 다시 脫아민化되어 피루브산 形態로 葉綠體에 들어간다. 그 以後는 NADP-ME型과 同一하게 進行되며 2 分子의 ATP 에너지를 使用해 PEP로 돌아온다. NAD-ME型 또한 1 分子의
農畜에는 2 分子의 ATP가 必要하다.
C4 植物의 進化
[
編輯
]
C4 植物은 白堊紀(藥 1億 3500萬 ~ 6500萬 年 前)에 처음 出現한 것으로 보이며 大氣 中의
濃度가 顯著하게 減少한 時期인 700萬 年 前에 顯著하게 增加하였다. 낮은
濃度에서 C3 植物보다 光呼吸이 적은 C4 植物 쪽이 生育에 유리한 境遇가 많기 때문에 C4 植物은 낮은
濃度에 適應했다는 假說도 있고 乾燥한 環境에서도 잘 자라기 때문에, 乾燥한 環境에 適應하여 進化했다고 하는 假說도 있다. 그러나 C4 植物은 進化의 起源이 여러 곳에서 由來하여 多元的으로 進化하고 있다. 속씨植物의 큰 두 갈래 植物群(單子葉植物, 雙子葉植物) 兩쪽 모두에서 C4植物을 볼 수 있기 때문에 속씨植物이 두 갈래의 무리로 나뉘기 前에 C4植物에 特異的으로 關聯된 遺傳子軍이 갖춰진 것으로 보고 있다. 實際로 벼와 같은 C3 植物에서 光合成에는 關與하지 않는 C4 植物의 것과 같은 PEPC, PPDK等의 遺傳子의 存在가 確認되고 있는데 C3 植物에서는 C4 植物과는 달리 遺傳子軍의 發現이 抑制되어 있는 것으로 보인다.
C3 植物은 高溫, 乾燥, 낮은
濃度, 窒素가 不足한 土壤 條件에서는 光呼吸이 쉽게 일어나
를 固定하기 힘들어진다. 하지만, C4 植物은 이러한 苛酷한 氣候條件에 適應하여
를 固定하기 위한 回路를 따로 가지게 되었다. 낮은 溫度에서는 別途의
固定回路가 에너지를 不必要하게 消耗하지만 乾燥한 條件에서는 生存에 바람직하게 된다. 作物 中 옥수수, 雜穀類가 C4 植物이며 벼나 밀은 C3植物이다. 雜草에도 C4 植物이 있는데 熱帶 地域에서는 C4 植物(雜草)李 C3 植物(作物)의 生長速度를 壓倒하기 때문에 雜草로 因한 被害 深刻한 問題가 되어 境遇에 따라서는 그 해 農事를 망치는 境遇까지 있다고 한다. 이 때문에 一部 科學者들은 主要 作物에 없거나 잠재되어 있는 C4 關聯 遺傳子를 活性化시켜 作物을 C4 植物로 만드는 硏究를 하고 있다.
같이 보기
[
編輯
]
各州
[
編輯
]
- ↑
同種異形(同種異形): 같은 種類임에도 形態가 다름
- ↑
獨逸語로 花冠(花冠), 꽃다발王冠이라는 意味