 染色體 末端을 保護하는 텔로미어
染色體 末端을 保護하는 텔로미어
텔로미어
(telomere) 또는 末端小粒 (末端小粒)은 染色體의 끝部分에 있는 染色 小粒으로 細胞의 壽命을 결정짓는 役割을 한다.
[1]
이것은 卽 細胞時計의 役割을 擔當하는
DNA
의 조각들이다. 텔로미어는 그리스어의 '끝'(τ?λο?, telos)과 '副尉'(μ?ρο?, meros)의 合成語다. 細胞分裂이 일어나는 동안에 染色體와 DNA를 複製하는 酵素는 染色體의 끝部分으로 複製를 繼續할 수 없다. 텔로미어가 없는 狀態로 細胞가 分裂된다면 細胞에 關한 情報가 들어있는 染色體의 끝部分이 消失될 것이다. 텔로미어는 染色體의 끝部分을 막고있는 分解되지 않는 緩衝地域이라 할 수 있다. 하지만 細胞가 分裂되면서 텔로미어는 消失되며 텔로머레이스라는 逆轉寫酵素에 依해 補充된다.
發見
[
編輯
]
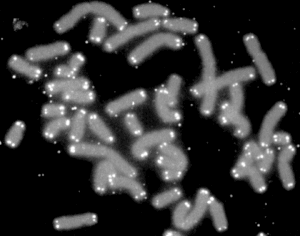 人間
染色體
(灰色) 끝 部分을 덮고 있는 텔로미어(흰色)
人間
染色體
(灰色) 끝 部分을 덮고 있는 텔로미어(흰色)
1970年代 初, 러시아 理論價 알렉세이 올로브니코프(Alexei Olovnikov)가 그 自身의 끝 部分을 完全하게 複製해내지 못하는 染色體들을 發見했다. 이 發見과 헤이플릭(Hayflick)의 槪念에 기초하여 DNA의 一部는 細胞의 壽命이 끝날 때까지 DNA 複製 時 繼續 消失될 것이라 假說을 세웠다.
[2]
[3]
그러나, 그의 豫想은 細胞 老化와 不滅化에 對해 硏究하는 一部 硏究者에게만 알려졌다.
[4]
1975年에서 1977年 사이에 엘리자베스 블랙번(Elizabeth Blackburn)는 單純히 染色體 末端을 構成하는 DNA 조각을 가지고 非凡한 텔로미어의 特性을 發見했다.
[5]
Blackburn과 Carol Greider, 그리고 Jack Szostak은 染色體가 어떻게 텔로미어와 텔로머레이스 酵素에 依해 保護되는지 밝혀 노벨賞(Physiology of Medicine)을 受賞했다.
[6]
그럼에도 不拘하고, 1970年代에는 텔로미어가 줄어드는 메커니즘이 普通 細胞 分裂 回數에 制限을 둔다는 認識은 全혀 없었을 뿐만 아니라, 이러한 現象이 細胞 老化에 큰 影響을 줄 것이라고 提示하는 硏究도 없었다. 또한, 그 메커니즘이 壽命을 制限 시킨다는 認識도 없었다.
[7]
[8]
以後, 生命工學 企業인 Geron이 細胞 老化에서 텔로미어 縮小에 어떻게 影響을 미치는지 밝히기 위해 RNA와 텔로머레이스에 對하여, 細胞 不滅化(cell immortalization) 狀態에서 텔로머레이스의 再活性化에 對해 持續的으로 投資를 했다.
[9]
텔로미어의 硏究 過程
[
編輯
]
細胞의 老化에 對해서 具體的으로 硏究한 Leonard Hayflick 博士는 1961年, 生物과 臟器에 따라서 細胞의 分裂 回數가 定해져 있고, 그 後에 細胞가 老化해 죽는다는 事實을 밝혀냈다.
[10]
Hayflick 博士는 胎兒의 細胞는 100番 程度 分裂하고, 老人의 細胞는 20~30番 程度 分裂한 後에 老化가 된다는 事實을 發見했다.
[11]
이를 헤이플릭 리미트(Hayflick Limit)라고 불린다. Hayflick 博士의 硏究에 依하면 고양이는 8番, 말은 20番, 人間은 60番 程度 細胞分裂을 할 수 있다고 한다. 그 後에 發見된 것이 바로 텔로미어이다. 1990年代 初가 되어서야 生物細胞學者들에 依해서 텔로미어가 染色體의 末端에 位置함이 밝혀졌다. 硏究가 繼續 進行된 結果, 샌프란시스코 캘리포니아 大學(UCSF)의 Elizabeth blackburn(61)敎授를 비롯하여 존스홉킨스 醫大 Carol Greider(48)과 하버드 醫大 Jack Szostak(57)은 텔로미어를 통해서 細胞의 老化 메커니즘을 糾明하였다.
[12]
이들은 2009年 노벨 生理學·醫學賞 受賞者로 選定되었다.
救助
[
編輯
]
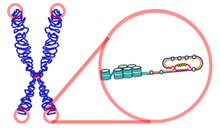 텔로미어의 T-loop 構造와 D-loop 救助
텔로미어의 T-loop 構造와 D-loop 救助
末端
[
編輯
]
텔로미어의 末端은 3`末端이 單一가닥으로 되어있는데, 이 部分은 t-loop(telomeric-loop)를 構成함으로써 安定된 構造를 形成한다. 이 안으로 묻히게 되는 構造를 'end-capping' 構造라 일컫는다. 二重가닥 中 튀어나온 한 가닥을 G-strand라 하는데, 이는 置換이 되어(displace) 'D-loop(displacement-loop)'를 形成하고 있다. 이러한 構造들은 染色體 末端 構造를 保護하기 위한 것으로 여겨진다. 그러나 텔로미어가 融合되면서 두 個의 動原體를 갖는 染色體(dicentric chromosome)를 만들 수 있고, 이는 窮極的 遺傳子 再配列을 일으켜 誘電體의 不安全性을 招來할 수 있다.
[13]
텔로미어-聯合蛋白質(telomere-associated proteins)
[
編輯
]
텔로미어 結合 蛋白質(telomere-binding proteins)
[
編輯
]
 쉘터린 複合體 救助
쉘터린 複合體 救助
現在까지 哺乳類에서 發見된 텔로미어 DNA에 鹽基序列 特異的으로, 直接的으로 結合할 수 있는 텔로미어 蛋白質은 TRF1, TRF2, POT1이다.
| 텔로미어 DNA 序列에 結合하는 蛋白質
|
| TRF1
|
텔로미어 길이 유지의 陰性的 調節(negatively regulated)
|
| TRF2
|
텔로미어 末端 保護에 重要한 調節 役割
|
| POT1
|
單一가닥 DNA 部分인 텔로미어 TTAGGG overhang에 結合
|
쉘터린(shelterin) 複合體
[
編輯
]
텔로미어에 直接的으로 結合하지는 못하지만(TRF1, TRF2, POT1과 달리) 텔로미어 蛋白質 複合體에 參與할 수 있는 蛋白質도 있는데 이를 텔로미어 聯合 蛋白質이라 한다. 위 言及한 세 蛋白質 包含하여 RAP1, TPP1, TIN2 等이 그 例다. 이들을 쉘터린 複合體라 일컫는다. 텔로미어에 쉘터린 複合體가 結合해 穩全한 텔로미어를 構成하면 텔로미어에서 DNA 修繕 期作이 作動하여 再組合을 일으키는 것을 막아준다. 또한, 텔로머레이스가 텔로미어로 接近하려 하는 것을 沮止한다. TIN2는 텔로미어 DNA 序列에 直接的으로 結合하지는 못하지만, 그 代身 다른 蛋白質인 TPP1과의 結合으로 TRF1 및 TRF2와 POT1을 이어주는 다리 役割을 한다. TPP1은 POT1과 POT1-TPP1 複合體를 形成한다(TPP1은 텔로미어 DNA 序列에 直接的으로 結合하지 못한다).
[14]
[15]
| 텔로미어 聯合 蛋白質(人間)
|
機能(推定)
|
텔로미어 結合 자리
|
| 二重가닥
|
單一가닥
|
| TRF1
|
텔로미어 길이 調節, 機能 維持
|
o
|
|
| TRF2
|
텔로미어 末端 保護, 텔로미어 길이 調節
|
o
|
|
| Ku70/80, DNA-PK 複合體
|
DNA 損傷 修繕, 텔로미어 길이와 構造 調節
|
o
|
|
| Tankyrase (TANK1, TANK2)
|
TRF1의 ADP 리보實話를 통한 텔로미어 길이 調節(positively regulate telomere length)
|
DNA에 直接 結合
못하고 TRF1과 上
號 作用함으로써
參與
|
|
| TIN2(TRF-interacting protein 2)
|
TRF1의 構造的 機能을 仲裁(negatively regulate telomere length)
|
TRF1과 相互 作用
|
|
| RAP1(repression and activation protein 1)
|
텔로미어 길이 調節(positively regulate telomere length)
|
TRF2와 相互 作用
|
|
| POT1
|
텔로미어 길이 調節, 텔로미어 末端 保護, 單一가닥 overhang(突出部分)部分의 保護
|
|
o
|
| hTERT
|
텔로머레이스의 觸媒 小段位(catalytic subunit)
|
|
|
| Ku
|
DNA 修繕(repair)
|
TRF1, TRF2와
相互作用
|
|
| MRN(MRE11/RAD50/NBS1)
|
細胞週期確認點(cell cycle checkpoint) 調節, DNA 修繕
|
TRF2와 相互 作用
|
|
| PinX1
|
텔로머레이스 抑制
|
TRF1, 텔로머레이
스와 相互 作用
|
|
TRF1이나 TIN2를 科發現 시킬 境遇 텔로미어 길이가 짧아진다.
열린 構造와 닫힌 構造
[
編輯
]
닫힌 構造 狀態의 텔로미어는 텔로머레이스가 텔로미어 길이를 신장 시키는 것을 妨害하고, 열린 構造 狀態의 텔로미어는 텔로머레이스와의 結合을 쉽게 하여 텔로미어 길이 增加를 誘導할 수 있다. 쉽게 말해 텔로미어가 닫힌 狀態여야 安定된 狀態라는 것이다. 쉘터린 救助는 텔로머레이스의 텔로미어에 對한 接近 可能性을 間接的으로 制限시켜 텔로미어 길이의 陰性的(negative) 調節에 關與한다. 따라서 쉘터린 蛋白質들은 '닫힌 텔로미어 救助' 또는 'cap의 形成(capping)'을 促進하고, 텔로미어 單一가닥 突出部分(3`-overhang)에 텔로머레이스가 接近하는 것을 防止한다.
[16]
性別에 따른 텔로미어 길이의 差異
[
編輯
]
女性의 텔로미어가 男性이 가지는 것보다 더 길다고 밝혀졌다. 서던 法(Southern blot)을 통해 이와 같은 結論을 導出해 냈으나, 이 發見은 아직 方法論的 差異가 있는 說明이 더 必要하다.
[17]
텔로미어의 作用
[
編輯
]
텔로미어의 길이는 種에 따라서 매우 다양하다. 酵母에서는 300~600個의 鹽基雙으로 이루어져있고, 人間의 境遇 數 킬로베이스(DNA 等 核酸 連鎖의 길이 單位)로 이루어져 있다. 人間의 境遇 텔로미어의 길이가 짧아짐에 따라서 細胞分裂을 막는 老化現象을 誘發할 수 있다. 텔로미어는 6個(人間의 境遇)의 特異的인 DNA 鹽基序列이 數百에서 數千 番 反復되며, 染色體의 末端에 位置하고 있어서 細胞가 分裂할 때 染色體가 分解되는 것을 막아준다. 텔로미어의 特徵을 살펴보면, 細胞가 한 番 分裂할 때마다 染色體 末端으로부터 50~200個의 텔로미어 DNA
뉴클레오타이드
를 잃어버린다. 텔로미어의 길이가 짧아질수록 細胞가 늙었다는 것을 意味한다. 그렇기 때문에 여러 次例 細胞分裂을 하면서 大部分의 텔로미어 DNA가 損失되면 細胞는 細胞分裂을 멈춘다.
텔로미어 鹽基序列
[
編輯
]
몇 가지 알려진 텔로미어의 鹽基序列
| 生物體
|
鹽基序列 (5'에서 3'을 向함 )
|
| 脊椎動物(vertebrate)
사람
생쥐
(Mice)
쥐
(Rats)
鳥類
|
TTAGGG
|
| 붉은빵곰팡이
|
TTAGGG
|
| 애기長대
|
TTTAGGG
|
| 클라미도모나스
|
TTTTAGGG
|
| 누에나방
|
TTAGG
|
| 酵母
|
TTAC(A)(C)G(1-8)
|
텔로머레이스
[
編輯
]

損失되는 텔로미어의 DNA를 復舊하는
酵素
가 存在하는데 그것을 텔로머레이스(末端小粒 複製酵素, telomerase)라고 한다. 이 酵素 德分에 細胞가 分裂해도 텔로미어의 길이를 어느 程度의 길이로 維持할 수 있다. 酵素가 지나치게 活性化되면 細胞가 繼續 分裂할 수 있다. 진핵 細胞로 이루어진 生物體에서 텔로머레이스가 活潑한 細胞는 所長 內部의 表皮細胞(
上皮細胞
),
骨髓
細胞,
癌
細胞 等이다. 所長 內部의 表皮細胞는 끊임없이 飮食物, 體液과 接觸하면서 傷處를 입거나 떨어져 나가곤 한다. 하지만, 活性化된 텔로머레이스에 依해서 細胞分裂이 持續的으로 일어나면서 傷處입거나 떨어져 나간 表皮細胞를 補充할 수 있는 것이다.
텔로머레이스는 텔로미어의 DNA 鹽基序列과 相補的인 鹽基雙을 가지는 RNA를 利用하여 텔로미어 DNA를 만들어내는 一種의
逆轉寫 酵素
를 만들어 낼 수 있다. (RNA의 相補的인 鹽基雙 : C(Cytosine)-G(Guanine), A(Adenine)-U(Uracil)) 例를 들어서 逆轉寫酵素의
RNA
의 鹽基序列이 AAUCCC이면 TTAGGG를 갖는 텔로미어 DNA鹽基序列이 만들어지는 것이다. 이와 같은 方法으로 텔로머레이스는 染色體 末端의 텔로미어의 길이가 짧아지지 않도록 해준다.
텔로미어의 縮小(細胞 老化)
[
編輯
]
 複製 될수록 줄어드는 DNA 二重 가닥. 仔細한 原理는 아래參照.
複製 될수록 줄어드는 DNA 二重 가닥. 仔細한 原理는 아래參照.
DNA 複製
가 DNA 末端에서부터 始作되지 않고 中央(複製 分岐點)에서 始作된다는 點,
DNA 重合酵素
는 DNA의 5`에서 3` 方向으로 移動한다는 點 때문에(DNA 複製가 5`에서 3`方向으로 複製 된다는 것을 勘案하면), 텔로미어는 漸次 部分的으로 짧아지게 된다.
先導 가닥에선 DNA 重合酵素가 5`에서 3`로 쭉 進行하기 때문에 相互 補完的인 DNA 가닥을 만들 수 있다. 그러나 遲延 가닥은 이와 같이 進行되지 않기 때문에 問題가 생긴다. 이 問題를 處理하기 위해서
프라이머
役割을 하는 짧은 RNA 조각 一部(RNA 프라이머)가 複製 分岐點 앞 部分에 붙는다. DNA 重合酵素는 그 部分에서부터 複製를 始作 시킬 수 있고, 複製 分岐點에서 끝으로 移動한다(5`에서 3`方向으로). 그러나 遲延 가닥에선 DNA 重合 酵素가 連續的으로 끊김 없이 5`에서 3`로 移動할 수 없기 때문에
오카자키 절편
이 만들어지게 된다. 더 많은 RNA 프라이머들이 DNA 가닥에 붙고 난 後에야 DNA 重合酵素들이 繼續해서 複製가 進行되도록 한다.
마지막 RNA 프라이머가 붙게되면 DNA 重合酵素, RNA 뉴클레이스, DNA 連結酵素가 RNA 彫刻 部分을 DNA로 變換시키고 오카자키 절편들의 사이를 메우기 위해 그 마지막 RNA 프라이머와 함께 온다. 그러나, RNA 部分(RNA 프라이머)을 DNA로 전환시키기 위해서는 RNA 프라이머의 앞部分에 또 다른 DNA 가닥(部分)이 있어야 한다. 그래야만 하는 理由가 末端의 RNA 프라이머가 除去된 後, 5` 쪽에는 dNTP를 重合할 3`?OH가 存在하지 않기 때문이다. 이것은 遲延 가닥 모든 部分에서 恒常 일어나는데 마지막 RNA 프라이머가 붙은 部分 끝에선 그 앞에 DNA部分이 없으므로 일어나지 않는다. 그러한 理由로, 이 境遇에 該當하는 RNA 프라이머 部分은 複製된 DNA에서 RNA部分을 分解하는 酵素에 依해 그냥 除去되어 버린다. 따라서 DNA 複製가 일어날 때 딸 細胞의 遲延가닥 5` 末端의 텔로미어는 持續的으로 漸次 조금씩 사라진다.
[18]
[19]
그러나 몇몇 試驗管에서 이루어진 硏究들은 텔로미어가 酸化스트레스(
oxidative stress
,
活性酸素
參照)에 매우 敏感하다는 것을 밝혔다. 또, 텔로미어의 結實(縮小)에 있어 스트레스 媒介 酸化的인 DNA 損傷(oxidative stress-mediated DNA damage)李 重要한 決定 要因이라는 證據도 있다.
[20]
Long Island Breast Cancer Study Project (LIBCSP)에서 硏究陣은 가장 짧은 텔로미어를 가지고 있고 베타 카로틴, 비타민 C와 E 攝取를 적게하는 女性들이 그렇지 않은 女性들보다 乳房癌에 걸릴 危險이 조금 더 크다고 밝혔다.
[21]
이러한 結果들을 통해 텔로미어의 縮小로 인한 癌 誘發 危險性이 DNA 損傷, 酸化 스트레스 같은 메커니즘들과 상호작용한다는 것을 類推해 볼 수 있다.
프라이머가 縮小되는 現象은 老化, 나이와 聯關된 疾病들, 死亡하는 것과 關聯이 있다고 豫測된다. 더 긴 텔로미어를 가진 生物이 짧은 텔로미어를 가지고 있는 生物보다 더 오래 살 수 있다는것은 Richard Cawthon가 2003年 밝혔지만,
[22]
짧은 텔로미어가 單純히 細胞의 나이를 標示하는지 老化에 直接的으로 寄與를 하는지는 밝혀진 바 없다.
[23]
老化가 進行되었다고 해서 모든 臟器나 機關들의 細胞에서 텔로미어가 一括的으로 短縮되지는 않는다. 또한 쥐의 텔로미어 길이(20~100 kb)가 우리의 것(5~15 kb)보다 김에도 不拘하고 오래 살지 못하는 것을 보면 더욱 그러하다.
에이즈 바이러스(HIV)와의 聯關性
[
編輯
]
에이즈 感染과 짧은 텔로미어의 聯關性은 에이즈 바이러스가 細胞, 生物學的 老化와 많은 나이와 關聯된 疾病의 影響에 間接的으로 影響을 주는 그 짧은 텔로미어에 影響을 준다고 提示했다. 實際로 에이즈 바이러스 感染軍과 非感染君을 比較했을 때, 非感染軍이 吸煙率과 알코올 攝取量이 높았음에도 不拘하고 바이러스 感染君의 텔로미어가 相當히 더 짧았다
[24]
吸煙
[
編輯
]
한 硏究에서 一生동안
담배
를 한番도 피운적 없는 사람과
吸煙
經驗이 있는 사람들의 텔로미어 길이에 對하여 統計를 내었다. 吸煙은 肺의 機能을 감소시켰고, 이것은 곧 긴 텔로미어의 縮小를 惹起했다. 한番도 吸煙을 하지 않은 사람과 比較했을 때, 짧은 텔로미어를 가지는 吸煙者들은 强制호기량(强制呼氣量)이 相當히 줄어들었다. 짧은 텔로미어를 가진 사람들은 吸煙으로 인한 肺 機能 減少 現象이 加速化되었다는 것이다.
[25]
肺 移植 手術
[
編輯
]
肺 移植 手術에서 寄贈者와 患者 텔로미어 길이의 斷面積인 特性은 以前에는 알려지지 않았다. 나중에야 獲得되고 태어날 때부터 타고나는 텔로미어 縮小 現象은 COPD(慢性 閉塞性 肺疾患)와 IPF(特發性 肺纖維症)를 包含한 末期 肺病에서 言及되었기 때문에, 한 硏究陣은 弊를 移植 받은 患者가 寄贈者 보다 더 짧은 텔로미어를 가질것이라고 假說을 세웠다.
108名을 對象으로 調査한 結果, 移植 手術을 받은 患者들의 텔로미어 길이가 寄贈者들에 비하여 相當히 짧았다. 이러한 現象은 寄贈者와 患者의 나이가 같을 때에도 發見되었다. 患者들의 텔로미어 길이 減少는 先天的인 現象이나 加速化된 縮小 때문일 것이다. 그러나 臟器 移植 以後의 寄贈者와 患者의 變化된 텔로미어 길이 役割은 밝혀지지 않았다.
[26]
延長
[
編輯
]
 헤이플릭 限界. 텔로미어가 줄어든다.
헤이플릭 限界. 텔로미어가 줄어든다.
制限된 細胞 分裂 現象은 Leonard Hayflick에 依하여 發見되었고, 이를
Hayflick limit
(헤이플릭 限界, 培養器 속에서 細胞가 生存하는 限界)라 일컫는다.
[27]
[28]
헤이플릭 限界 發見을 始發點으로, 텔로미어가 縮小되는 現象을 발 묶는 아이디어는 後에 Michael D. West가 創立한 Geron Corporation의 硏究陣이 發見했다.
[29]
텔로머레이스
(텔로미어 신장 酵素) 觸媒 物質을 複製하여 特定 條件에서의 텔로머레이스 作用이 텔로미어 縮小를 防止하여 人間의 細胞를 不滅로 만들 수 있는지 確認하는 테스트를 했다. 1998年에 "텔로머레이스가 細胞 壽命을 延長시킬수 있다"라는 結果는 公開的으로 發表 되었고, 只今은 人體 細胞를 不滅化 시킬 수 있을 것으로 期待된다.
[30]
텔로머레이스를 活性化시킴으로써 텔로미어 縮小를 逆으로 작용시켜 老化가 正常보다 느리게 進行되도록 할 수 있다는 것은 분명해지고 있다. 이는 헤이플릭 限界를 延長 시키므로 人間의 壽命은 그에 따라 늘어날 것이다. 또한 藥물, 遺傳子 治療, 無氣力함이나 東面과 같이 物質 代謝를 抑制하는 이 세 가지 方法을 통해 텔로미어 縮小를 反轉시킬 수 있는 것으로 提示되었다. 아직까지 人體에선 앞서 提示된 方法들이 效果가 있다고 直接的으로 밝혀진 바는 없으나, 텔로미어 縮小가 逆으로 作用하는 冬眠 狀態에서 老化가 抑制된다는 것(Turbill, et al. 2012 & 2013)과 冬眠하는 것이 壽命을 延長시킨다는 것(Lyman et al. 1981)은 立證되었다. 또 實驗用 쥐와
[31]
[32]
예쁜꼬마선충
을
[33]
利用해 텔로미어 연장이 그 個體의 老化 信號들을 逆으로 작용시킨 實驗을 해냈다. 그러나 텔로머레이서가 作用하는 긴 텔로미어는
癌細胞
가 될 確率이 높다고 假說이 세워졌다(e.g. Weinstein and Ciszek, 2002). 또, 그 逆으로 긴 텔로미어는 癌으로부터 保護를 하기도 하는데 짧은 텔로미어들은 癌과 密接하기 때문이다. 그리고 긴 텔로미어가 에너지 消費量을 增大시킬 수 있다고 提示되었다.
[34]
텔로미어를 延長시키는 技術은 生物 醫學 治療 技術에 使用될 수 있는 充分히 많은 量의 健康하고, 癌化되지 않은 哺乳類의 細胞들을 培養하는 것을 可能하게 하기 때문에 組織工學에 도움이 될 것으로 展望된다.
長壽(長壽)韓 바닷새에 關한 두 硏究는 텔로미어의 役割이 아직까지 完璧하게 밝혀지지 않았다는 것을 보여준다. 2003年, 科學者들은 바다제비(
Leach's storm-petrel
,
Oceanodroma leucorhoa
)의 텔로미어가 生活 年齡에 따라 길어지는 現象을 觀察했다.
[35]
이러한 텔로미어 길이의 變化는 처음으로 觀察되었다. 그리고 2006年, Juola 外 몇名
[36]
이 또 다른, 壽命이 긴 바닷새 종인 큰軍艦鳥의 텔로미어 길이는 大略 最小限 40年 동안(前 生을 걸쳐) 持續的으로 줄어들었으나, 나이가 듦에 따라 그 速度가 確然하게 줄어들고 이러한 텔로미어 길이의 減少率은 各 새 個體에 따라 매우 다양했다. 硏究陣은 이 鐘(軍艦새와 그의 親戚種들)에서 텔로미어의 길이는 한 새의 나이를 判別하기에 充分하지 않은 것이라 結論내렸다. 처음 믿어졌던 것 보다도 텔로미어 길이의 習性은 훨씬 더 다양한 것으로 期待된다.
텔로미어의 길이와 壽命에 對한 疑問點
[
編輯
]
고메즈 外 硏究陣(Gomes et al)李 哺乳類 動物 텔로미어의 比較 生物學的 硏究에서 서로 다른 哺乳類 種의 텔로미어 길이는 壽命과 直接的이진 않다고 밝혔으며 壽命에 對한 텔로미어 길이의 影響이 如前히 疑問이라고 結論을 내렸다. 헤리스 外 硏究陣(Harris et al)은 "人間 텔로미어 길이는 認知的 能力과 身體 能力에 對한 老化의 重要한 生物指標다"라는 證據를 찾았다. 길리와 블랙번(Gilley and Blackburn)은 짚신벌레를 利用하여 細胞 老衰期가 텔로미어 縮小에 依한 것인지를 알아보기 위해 實驗했으나, 細胞 老衰期 동안 텔로미어는 줄어들지 않았다.
[37]
生活 習慣과의 聯關性
[
編輯
]
캘리포니아 大學에서 推進된 한 準備 調査는 初期 前立腺 癌 診斷을 받은 35名을 對象으로 했고 그 中 10名의 生活 方式을 바꾸게 했다. 菜食(과일, 菜蔬, 특별한 化學 處理가 안된 穀物, 低脂肪, 精製된 糖質), 適當한 運動(1週日에 6日 30分씩 걷기), 스트레스 減少시키기(요가 스트레칭, 呼吸, 冥想), 週間 그룹 支援이 그 方式들이다. 그 外 25名 參加者들과 比較했을 때, 生活 習慣을 바꾼 그룹에서 大略 10%의 더 긴 텔로미어가 觀察되었다. 게다가 얼마나 그 要請에 應했는지(얼마나 熱心히 自身의 生活 方式을 바꾸었는지)에 따라서도 더욱 相當한 差異가 있었다. 2014年, "健康을 爲해 일어서라. 앉아서 하는 行動을 避하는 것은 當身의 텔로미어를 延長시킬 것이다."란 標題를 지닌 한 硏究는 多少 矛盾되는 結果를 提示했는데, 다음과 같이 結論을 내렸다. "仲裁 그룹에서 텔로미어 길이의 變化와 運動하는데 보낸 時間 變化 사이에 否定的인 聯關性이 있었다(rho=-0.39, p=0.07). 그런데, 仲裁 그룹에서 텔로미어 延長은 앉아있는 時間을 줄이는 것과 相當히 聯關이 있었다(rho=-0.68, p=0.02)"
텔로미어와 癌細胞
[
編輯
]
癌細胞
는 끊임없이 細胞分裂을 하기 위해서 텔로미어 DNA의 길이를 維持할 수 있는 메커니즘이 必要하다. 텔로미어의 길이를 延長하거나 維持하는 메커니즘은 細胞 單位의 끊임없는 增殖을 하기 위해서 꼭 必要한 過程이다. 텔로미어를 延長하기 위해 必要한 酵素인 텔로머레이스는 腫瘍의 90%에서 活性化되어 있다. 그렇기 때문에 癌細胞에서는 다른 體細胞에 比해서 壽命이 길어진다. 이를 통해 癌細胞에 있는 텔로머레이스의 機能을 抑制하거나 癌細胞의 텔로미어 DNA를 除去하면, 癌細胞의 細胞分裂을 막을 수 있다.
p53이나 pRb와 같은 腫瘍抑制遺傳子가 存在하면 細胞老化가 일어나고 그렇지 않으면 어느程度 老化가 進行된 體細胞가 癌細胞로 變할 수 있다.
[38]
[39]
最近 實驗및 免疫學 敎授이자 이 大學의 벡스터 줄기細胞 生物學硏究所(Baxter Laboratory for Stem Cell Biology) 이사인 헬렌 블라우(Helen Blau) 博士는 人間 텔로미어 길이를 1,000 뉴클레오타이드 길이만큼 늘이는 方法(TERT 시퀀스가 코딩된 mRNA를 利用하여)을 發見했다. 이는 사람 壽命으로 數千年에 該當한다.
[40]
기타
[
編輯
]
2009年
노벨賞
數上位는 텔로미어와 텔로머레이스의 硏究로
잭 쇼스택
外 2名에게
노벨 生理學·醫學賞
을 授與했다.
같이 보기
[
編輯
]
各州
[
編輯
]
- ↑
우리말샘 - 末端小粒 等
- ↑
Olovnikov, Alexei M (1971). “Principle of marginotomy in template synthesis of polynucleotides (Принцип маргинотомии в матричном синтезе полинуклеотидов)”. 《Doklady Akademii Nauk SSSR》.
- ↑
Olovnikov AM (September 1973). “A theory of marginotomy. The incomplete copying of template margin in enzymic synthesis of polynucleotides and biological significance of the phenomenon”. 《J. Theor. Biol.》.
- ↑
“No Nobel physiology and medicine award for Russian gerontologist Aleksey Olovnikov”
. 2009年 10月 21日. 2016年 3月 11日에
原本 文書
에서 保存된 文書
. 2016年 12月 8日에 確認함
.
- ↑
Blackburn AM (March 1978). “A tandemly repeated sequence at the termini of the extrachromosomal ribosomal RNA genes in Tetrahymena”. 《J. Mol. Biol.》.
- ↑
“The 2009 Nobel Prize in Physiology or Medicine - Press Release”
. Nobelprize.org. 2009年 10月 5日
. 2012年 6月 12日에 確認함
.
- ↑
Harrison's Principles of Internal Medicine, Ch. 69, Cancer cell biology and angiogenesis, Robert G. Fenton and Dan L. Longo, p. 454.
- ↑
“Portfolio”
.
- ↑
“Unravelling the secret of ageing”
. 《COSMOS: The Science of Everything》. 2009年 10月 5日. 2015年 1月 14日에
原本 文書
에서 保存된 文書.
- ↑
Douglas E. Crews.
《Human Senescence: Evolutionary and Biocultural Perspectives》
. Cambridge University Press. 49쪽
. 2016年 12月 8日에 確認함
.
- ↑
Herman Todorov, Robert Nadler, Igor N. Todorov (2000).
《Public Enemy Number 1--stress: A Practical Guide to the Effects of Stress and Nutrition on the Aging Process and Life Extension》
. Nova Publishers. 18쪽
. 2016年 12月 8日에 確認함
.
- ↑
ELIZABETH H. BLACKBURN (1991年 4月 18日).
“Structure and function of telomeres”
. 《nature》. nature.
- ↑
오상진. 《老化의 生物學:分子生物學的 原因에 對하여》. 探究當. 85-86쪽.
- ↑
Blasco, Maria (2010年 6月 21日).
“Role of shelterin in cancer and aging”
. 《Aging Cell》.
- ↑
오상진. 《老化의 生物學:分子生物學的 原因에 對하여》. 探究當. 91쪽.
- ↑
오상진. 《老化의 生物學》. 探究當.
- ↑
“Gender and telomere length: systematic review and meta-analysis.”
. 《NCBI》.
- ↑
“Telomerase”
. 《PediaPress》.
- ↑
K. Lenhard Rudolph (2007. 11. 7).
《Telomeres and Telomerase in Aging, Disease, and Cancer: Molecular Mechanisms of Adult Stem Cell Ageing》
. Springer Science & Business Media. 212쪽.
- ↑
Richter, T; von Zglinicki, T (2007). “A continuous correlation between oxidative stress and telomere shortening in fibroblasts”. 《Exp Gerontol》
42
(11): 1039?1042.
doi
:
10.1016/j.exger.2007.08.005
.
PMID
17869047
.
- ↑
Shen, J (Apr 2009). “Telomere length, oxidative damage, antioxidants and breast cancer risk”. 《Int J Cancer》
124
(7): 1637?43.
doi
:
10.1002/ijc.24105
.
- ↑
Richard M Cawthon MD, Ken R Smith PhD, Elizabeth O'Brien PhD, Anna Sivatchenko MD, Richard A Kerber PhD (01 February 2003).
“Association between telomere length in blood and mortality in people aged 60 years or older”
. 《The Lancet》.
- ↑
Karen Anne Mather, Anthony Francis Jorm, Ruth Adeline Parslow and Helen Christensen (2010年).
“Is Telomere Length a Biomarker of Aging? A Review”
. 《Oxford Journals》.
- ↑
Elizabeth Auld (2016年 9月 21日).
“HIV Infection Is Associated with Shortened Telomere Length in Ugandans with Suspected Tuberculosis”
. 《journals.plos.org》.
- ↑
Pascal Andujar, MD-PhD 1 , Dominique Courbon, MSc 2 , Emilie Bizard, Student 1 , Elisabeth Marcos, PhD 1 , Serge Adnot, MD 1 , Pascal Demoly, MD PhD 3 , Isabelle Pin, MD PhD 4 , Gabriel Thabut, MD, PhD, Prof 5 , Jorge Boczkowski, MD PhD 1 , Benedicte Leynaert, PhD 2 ,.
“Associations Between Smoking, Telomere Length and Lung Function Decline: Findings from a Population-Based Longitudinal Study”
. 《ATS Journals》.
- ↑
Courtwright AM, Fried S, Villalba JA, Moniodis A, Guleria I, Wood I, Milford E, Mallidi HH, Hunninghake GM, Raby BA, Agarwal S, Camp PC, Rosas IO, Goldberg HJ, El-Chemaly S. (2016 Sep).
“Association of Donor and Recipient Telomere Length with Clinical Outcomes following Lung Transplantation.”
.
- ↑
Hayflick L, Moorhead PS (1961). “The serial cultivation of human diploid cell strains”. 《Exp Cell Res》
25
(3): 585?621.
doi
:
10.1016/0014-4827(61)90192-6
.
PMID
13905658
.
- ↑
“The limited in vitro lifetime of human diploid cell strains”. 《Exp. Cell Res.》
37
(3): 614?636. 1965.
doi
:
10.1016/0014-4827(65)90211-9
.
PMID
14315085
.
- ↑
Feng J, Funk WD, Wang SS, Weinrich SL, Avilion AA, Chiu CP, Adams RR, Chang E, Allsopp RC, Yu J (September 1995). “The RNA component of human telomerase”. 《Science》
269
(5228): 1236?41.
doi
:
10.1126/science.7544491
.
PMID
7544491
.
- ↑
Bodnar, A.G. (1998). “Extension of life-span by introduction of telomerase into normal human cells”. 《Science》
279
(5349): 349?352.
doi
:
10.1126/science.279.5349.349
.
- ↑
Sample, Ian (2010年 11月 28日).
“Harvard scientists reverse the ageing process in mice ? now for humans”
. The Guardian.
- ↑
Jaskelioff, Mariela.
“Telomerase reactivation reverses tissue degeneration in aged telomerase-deficient mice”
. 《Nature》
469
(7328): 102?106.
doi
:
10.1038/nature09603
.
PMC
3057569
.
PMID
21113150
.
- ↑
Joeng KS, Song EJ, Lee KJ, Lee J (2004). “Long lifespan in worms with long telomeric DNA”. 《Nature Genetics》
36
(6): 607?11.
doi
:
10.1038/ng1356
.
PMID
15122256
.
- ↑
Eisenberg DTA (2011). “An evolutionary review of human telomere biology: The thrifty telomere hypothesis and notes on potential adaptive paternal effects”. 《American Journal of Human Biology》
23
(2): 149?167.
doi
:
10.1002/ajhb.21127
.
PMID
21319244
.
- ↑
Nakagawa S, Gemmell NJ, Burke T (September 2004). “Measuring vertebrate telomeres: applications and limitations”. 《Mol. Ecol.》
13
(9): 2523?33.
doi
:
10.1111/j.1365-294X.2004.02291.x
.
PMID
15315667
.
- ↑
Juola, Frans A/Haussmann, Mark F/ Dearborn, Donald C/Vleck, Carol M (2006). “Telomere shortening in a long-lived marine bird: Cross-sectional analysis and test of an aging tool”. 《
The Auk
》
123
(3): 775.
doi
:
10.1642/0004-8038(2006)123[775:TSIALM]2.0.CO;2
.
ISSN
0004-8038
.
- ↑
Nuno M. V. Gomes,1,2 Oliver A. Ryder,3 Marlys L. Houck,3 Suellen J. Charter,3 William Walker,1,* Nicholas R. Forsyth,4 Steven N. Austad,5 Chris Venditt,6,† Mark Pagel,6,7 Jerry W Shay,1 and Woodring E. Wright1 (2011 Jun 1).
“Comparative biology of mammalian telomeres: hypotheses on ancestral states and the roles of telomeres in longevity determination”
. 《NCBI》.
- ↑
Keiko Hiyama (2009. 3. 18).
《Telomeres and Telomerase in Cancer》
. Springer Science & Business Media. 51-52쪽.
- ↑
오상진. 《老化의 生物學:分子生物學的 原因에 對하여》. 探究當. 64-73, 103-106쪽.
- ↑
John Ramunas*,1, Eduard Yakubov†,1,2, Jennifer J. Brady*, Stephane Y. Corbel*, Colin Holbrook*, Moritz Brandt*, Jonathan Stein‡, Juan G. Santiago§, John P. Cooke†,2 and Helen M. Blau*,3 (2015年 1月 22日).
“Transient delivery of modified mRNA encoding TERT rapidly extends telomeres in human cells”
. 《fasebj.org》.
參考 文獻
[
編輯
]
|
|---|
| 歷史
| | |
|---|
| 下位 分野
| |
|---|
| 生物學的 槪念
| |
|---|
| 一般 槪念
| |
|---|
基本 技術
및 道具
| |
|---|
| 應用
| |
|---|
| 學制 간 分野
| |
|---|
| 目錄
| |
|---|
|