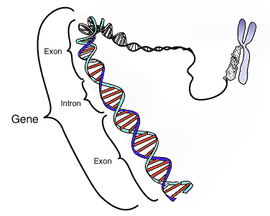
 遺傳子에서 인트론과 엑손의 位置 模式圖
遺傳子에서 인트론과 엑손의 位置 模式圖
진핵細胞
의 DNA에는
原核細胞
의 DNA와 달리 實際로 蛋白質을 合成하는 部分과 아무런 意味가 없는 部分이 섞여서 存在하고 있다. 이때 意味없는 部分을
인트론
(Intron)이라 한다.
따라서 眞核細胞에서는 DNA에서 mRNA를 만드는
戰死
(transcription) 過程에서 意味있는 部分인
엑손
끼리 잘라 잇는 過程을 거치게 된다.
인트론은 蛋白質을 만드는 데 關與하지 않는
DNA
部分으로 戰士過程에서 pre-mRNA를 만드는 데 使用되지만 matured mRNA를 만드는 데 쓰이지 않고 잘려진다. 인트론은
眞核生物
에서 흔하게 볼 수 있지만 原核生物에서는 그렇지 못하고 tRNA, rRNA만 存在한다. matured mRNA에 남아있는 DNA戰士部分을 엑손(exon)이라 한다. 인트론의 個數와 길이는 種(種)間에서뿐만 아니라 鐘 內部에서도 다르다.
Takifugu rubripes
라는 복魚는 인트론이 거의 없지만 哺乳類나 開花植物의 境遇 엑손의 길이보다 길고 많은 數의 인트론을 가지고 있다.
槪要
[
編輯
]
 pre-mRNA에서 mRNA 切斷 過程 圖式
pre-mRNA에서 mRNA 切斷 過程 圖式
한 遺傳子(gene)에서 여러 가지 蛋白質을 生産하기 위해서 인트론은 遺傳子의
代替揷入
(alternative splicing)을 許容하기도 한다. 代替揷入을 包含한 mRNA의 切斷은 다양한 信號分子에 依해 遂行된다. 예전에는 쓰인 적이 있지만 現在에는 쓰이지 않는 오래된 코드를 包含하기도 한다.
인트론의 大部分의 序列들은 알려진 機能이 없는 쓰레기 DNA라고 여겼지만 짧은 鹽基序列의 反復은 效果的인 mRNA切斷에 影響을 미친다는 것이 밝혀졌다. 인트론 切斷의 向上(intronic splicing enhancers)에 對한 正確한 過程은 알려져 있지 않지만 戰死過程에서 스플라이오솜(spliceosome)을 安定化시키며 蛋白質과 結合하는 部位로 作用할 것으로 보인다.
인트론의 發見으로 1993年 Phillip Allen Sharp와 Richard J. Roberts는 노벨 醫學賞을 受賞했다.
I 그룹과 II 그룹 인트론은 스스로 잘려지는 인트론이다. 傳寫된 primary RNA를 絶斷을 觸媒하는 리보自任(ribozyme)으로 불리며 RNA의 觸媒劑로서의 機能이 發見된 데 意義가 있다. 스스로 잘려지는 인트론의 發見者 Thomas Cech는 Sidney Altman과 1989年 노벨化學賞을 共同受賞하게 되었다.
인트론의 進化
[
編輯
]
인트론의 起源은
액틴
과 같은 잘 保存된 유전자군에서 活潑하게 硏究되었다. 인트론의 進化的 起源에 對해서는 2가지 有力한 理論(初期모델, 後記모델)이 있다.
初期 인트론 모델에서는 祖上의 遺傳子가 많은 數의 인트론을 包含했으나 進化를 거치고 종이 分化되면서 消失되거나 달라졌다(서로 다른 種의 類似한 機能의 遺傳子에서 비슷한 인트론 樣式을 보이기도 한다.)고 主張한다.
後記 모델에서는 새로운 인트론의 導入을 위해 어떠한 事前措置를 한다고 假定하고 이 때문에 特定 遺傳子 變異의 같은 場所에서 類似한 인트론이 나타난다고 主張한다. 全혀 다른 2個의 種에서 같은 인트론 類型을 나타나는
收斂進化
現象이 나타날 수 있다고 主張한다.
인트론 엑손의 構造變化는 漸進的이고 週期的이며 코딩順序의 進化와는 別個로 일어난다.
參考 文獻
[
編輯
]
- Walter Gilbert
(1978. 2. 9.) "Why Genes In Pieces?"
Nature
271
(5645):501.
外部 링크
[
編輯
]
