A
bakteriumok
(Bacteria)
egysejt?, tobbnyire nehany
mikrometeres
sejtes
mikroorganizmusok
. Valtozatos megjelenes?ek: sejtjeik gomb, palcika, csavart alakuak lehetnek. A
mikrobiologia
egyik aga, a
bakteriologia
foglalkozik a bakteriumok tudomanyos es elettani vizsgalataval.
A
Fold
minden el?helyen megtalalhatoak a bakteriumok: a vizben, a szarazfoldon vagy a leveg?ben, meg melytengeri h?forrasokban es nuklearis hulladekban is.
[2]
Egy gramm talaj kb. 40 millio, egy milliliter felszini viz egymillio bakteriumsejtet tartalmaz.
A
Foldon
pedig osszesen mintegy 5
kvintillio
(5 × 10
30
) bakterium elhet.
[3]
A bakteriumok alapvet? szerepet toltenek be a
bioszfera
anyagforgalmaban, mint peldaul a legkori
nitrogen
megkoteseben. Ennek ellenere a bakteriumfajok nagy reszet nem ismerjuk: a bakteriumtorzsek fele rendelkezik csak olyan fajokkal, amelyek laboratoriumi korulmenyek kozott tenyeszthet?k.
[4]
Valamivel tobb bakterium van az emberi testben, mint emberi sejt. A legtobb bakterium a b?r felszinen es az emeszt?rendszerben talalhato.
[5]
A bakteriumok nagy resze artalmatlan vagy hasznos, de akad nehany
fert?z? megbetegedest
kivalto
patogen
(korokozo) bakterium is, mint peldaul a
kolera
,
szifilisz
,
lepfene
,
lepra
vagy a
pestis
korokozoja. Gyakori es sulyos bakterialis megbetegedes a
tuberkulozis
(TBC), amely evente ketmillio embert ol meg nagyreszt
Afrikaban
, a
Szaharatol
delre es? teruleteken.
A fejlett orszagokban
antibiotikumokat
hasznalnak a fert?zesek lekuzdesere. Ezek tulzasba vitt hasznalata, kulonoskeppen pedig a bakteriumok szeles korere hato antibiotikumok kiterjedt hasznalata eredmenyekent egyre tobb
antibiotikumellenallo
tipus fejl?dott ki. Ennek egy specialis esete figyelhet? meg a
Clostridium difficile
bakteriumnal. Az
antibiotikum-ellenallas
elterjedesehez hozzajarult ezeknek a gyogyszereknek a helytelen hasznalata, az orvosi el?iras pontos betartasanak elhanyagolasa (
lasd lejjebb
).
Az iparban a szennyviztisztitasban, a tejtermekek gyartasaban, az antibiotikumok es mas szerves anyagok el?allitasaban hasznalnak bakteriumokat.
[6]
A bakteriumok
prokariota
szervezetek, tehat szemben az allatokkal es mas
eukariotakkal
, nincs
sejtmagjuk
es mas
membrannal
hatarolt
sejtszervecskejuk
. Ambar hagyomanyosan bakteriumnak neveznek minden prokariotat, a tudomanyos nevezektan az utobbi par evben megvaltozott, miutan molekularis biologiai modszerekkel a prokariotakat sikerult ket alapvet?en elter? felepites? es
szarmazasu
csoportra kuloniteni. Ez a ket
domen
az
Archeak
es a
Bacteria
.
[7]
A bakteriologia tortenete
[
szerkesztes
]
 Anton van Leeuwenhoek
,
mikroszkopja
segitsegevel el?szor figyelt meg bakteriumot
Anton van Leeuwenhoek
,
mikroszkopja
segitsegevel el?szor figyelt meg bakteriumot
Az els? bakteriumokat
Anton van Leeuwenhoek
[8]
holland
termeszettudos pillantotta meg
1674
-ben, egy sajat maga altal keszitett egylencses, ketszazszoros nagyitasra kepes
mikroszkopban
. Megfigyeleseit a Kiralyi Tarsasaghoz irt leveleiben publikalta.
[9]
[10]
Maga a
bakterium
elnevezes a
gorog
βακτηριον szobol szarmazik, melynek jelentese ?kis palca”;
[11]
a nevet
Christian Gottfried Ehrenberg
javaslatara
1828
-ban vezettek be.
A 19. szazad masodik feleben
Louis Pasteur
bizonyitotta a mikrobak szerepet az
erjedessel
,
rothadassal
es
fert?zesekkel
kapcsolatban. Pasteur nyoman
Joseph Lister
angol sebesz
1865
-ben felismerte, hogy a
sebfert?zes
okozoi is bakteriumok es orvosi m?szereit
karbolsavval
sterilizalta
. Ugyanebben az evben halt meg
Semmelweis Ignac
magyar szulesz-n?gyogyasz, aki
1847
-ben allati eredet? ?bomlott szerves anyag” nyilt serulesekre valo atvitelevel magyarazta, es klormeszes kezfert?tlenitessel akadalyozta meg a
gyermekagyi laz
(
Streptococcus
sebfert?zes) kialakulasat.
Robert Koch
19. szazadi
nemet
orvosbiologus
nevehez f?z?dik szamos bakteriumfaj azonositasa. Kulonboz? laboratoriumi technikai (peldaul a
lemeztenyeszet
) segitsegevel elkulonitette es azonositotta a
tuberkulozis
,
lepfene
es
kolera
korokozojat. A tuberkulozissal vegzett kutatasaiert Koch
1905
-ben
Nobel-dijat
kapott.
[12]
A
Koch-fele posztulatumok
? a betegsegek mikrobialis eredetere vonatkozo kovetelmenyek ? ma is hasznalatban vannak.
[13]
Habar mar a 19. szazadban ismert volt, hogy szamos betegseget bakteriumok okoznak, sokaig nem sikerult hatasos antibakterialis kezelest kidolgozni.
[14]
1910
-ben
Paul Ehrlich
fejlesztette ki az els?
antibiotikumot
. A
szifilisz
korokozojat, a
Treponema pallidum
nev?
spirochaetat
szelektiven fest? anyagban cserelt ki komponenseket oly modon, hogy az uj keverek a patogent szelektiven elpusztitotta.
[15]
Ehrlich szinten Nobel-dijat kapott az
immunologia
teruleten vegzett munkajaert, es elen jart a kulonboz? bakteriumok kimutatasara es azonositasara hasznalt festesi eljarasok kidolgozasaban. Az ? munkai kepeztek alapjat a
Gram-festesnek
es a Ziehl?Neelsen-festesnek is.
[16]
A bakteriumok tanulmanyozasaban jelent?s lepes volt
Carl Woese
azon felismerese
1977
-ben, hogy az
Archaeak
a bakteriumoktol elter? evolucios vonalat kepviselnek.
[17]
Ez az uj filogenetikus osztalyozas a 16S
riboszomalis
RNS
szekvenalasan alapult, es a prokariotakat ket evolucios domenre osztotta, kialakitva igy a 3 domenes rendszert.
[18]
Eredet es evolucio
[
szerkesztes
]
A mai bakteriumok ?sei egysejt? mikroorganizmusok, a Fold els? eletformai voltak, melyek 4 milliard evvel ezel?tt eltek. Mintegy 3 milliard eve az osszes el?leny mikroszkopikus meret? volt, a bakteriumok es Archaea domen ?si kepvisel?i voltak az elet dominans formai.
[19]
[20]
Habar leteznek bakterialis
kovuletek
, mint peldaul a
Sztromatolitok
, a jellegzetes morfologiai jegyek hianya nem teszi lehet?ve, hogy a bakterialis evoluciot, vagy egy bizonyos bakteriumfaj eredetet rajtuk keresztul lehessen tanulmanyozni. A genetika azonban lehet?ve teszi a bakterialis
torzsfejl?des
rekonstrualasat, es ezek a kutatasok arra utalnak, hogy a bakteriumok az Archaea vonaltol elvalva kezdtek el kulon uton fejl?dni.
[21]
A bakteriumok es az Archaea utolso kozos ?sei valoszin?leg azok a termofil szervezetek lehettek, melyek 2,5?3,2 milliard evvel ezel?tt eltek.
[22]
[23]
A masodik nagy evolucios szetvalasban, az archeak es az eukariotak szetvalasaban is szerepet jatszottak a bakteriumok. Az eukariotak akkor jelentek meg, amikor ?si bakteriumok
endoszimbiozisra
leptek az eukariota sejtek ?seivel, melyek maguk is feltehet?en az Archea csoport tagjai voltak.
[24]
Ennek soran az ?si forma bekebelezett egy alfa-proteobakteriumot (melyb?l kes?bb a
mitokondrium
lett) es egy cianobakteriumszer? organizmust (melyb?l kes?bb a
szintest
lett).
[25]
[26]
Ezt az un.
endoszimbionta-elmeletet
Lynn Margulis
(
1938
?) amerikai kutato
1967
-ben publikalta el?szor.
[27]
Margulis szerint a bekebelezett kisebb prokariota sejtek tovabb eltek a sejten belul, es az egyutteles sikeres sejtkapcsolatnak bizonyult. Az elmelet bizonyiteka lehet az, hogy a mitokondrium es a szintest bakterialis meret?; sajat orokit?anyaggal rendelkeznek, ami a prokariotakhoz hasonloan kor alaku
DNS
; sajat
enzimatikus
apparatussal rendelkeznek es
osztodasuk
a sejt osztodasatol fuggetlen.
 A bakteriumok sejtjeinek felepitese es elrendez?dese valtozatos kepet mutat.
A bakteriumok sejtjeinek felepitese es elrendez?dese valtozatos kepet mutat.
A bakteriumok alakja es merete nagyon valtozatos kepet mutat. A bakteriumsejtek az eukariota sejteknel kb. 10-szer kisebbek, leggyakrabban 0,5?5 mikrometer a hosszusaguk. Azonban akad nehany faj, mint peldaul a
Thiomargarita namibiensis
es a
Epulopiscium fishelsoni
,
melyek akar a fel
millimeteres
nagysagot is elerik, es szabad szemmel is lathatoak.
[28]
A legkisebb bakteriumok a
Mycoplasma
nemzetsegbe tartozo fajok; mindossze 0,3 mikrometeres meretuk megegyezik a legnagyobb
virusok
meretevel.
[29]
A legtobb bakteriumfaj gomb vagy palcika alaku. A gomb alakuak masik neve coccus a gorog
kokkos
szo utan, mely magot jelent. A palcika alakuak masik neve bacilus, a latin
baculus,
palca szobol szarmaztatva. Tipikus kepvisel?juk a kolibacilus
(
Escherichia coli
).
Nehany palcika alaku bakterium hajlott vessz? alaku (mas neven komma vagy vibrio alak), mint a koleravibrio
(
Vibrio cholerae
).
A spirillumok merev csavar alaku bakteriumok. A dugohuzo alaku, hosszu es nagyon vekony
spirochaetak
sejtfala nem merev, ezert mozgas kozben elhajolnak. Keves
tetraeder
vagy
kocka
alaku fajt is ismernek.
[30]
A bakteriumok alakjat a bakterialis
sejtfal
es a
citoszkeleton
(sejtvaz) hatarozza meg. Az alak alapvet?en befolyasolja, hogy a bakterium hogyan tud taplalekot szerezni, letapadni, folyadekban uszni, vagy tamadoi el?l elmenekulni.
[31]
[32]
A sejtfallal eredetileg rendelkez?, de azt elveszitett,
L-forma
bakteriumegyedek ezert a kiindulasi bakterium alakjatol fuggetlenul
gomb
vagy
szferoid
alakuak.
Szamos bakteriumfaj egyetlen sejtkent eli le eletet, masok jellegzetes mintazatot alkotva tarsulnak es csoportokat vagy telepeket kepeznek egymassal: a
Neisseria
fajok parokat (diploidokat) kepeznek, a
Streptococcusok
lancot alkotnak, a
Staphylococcusok
sz?l?furtszer?en csoportosulnak. A bakteriumok fonalszer?en megnyulhatnak, mint peldaul az
Actinobacteria
(sugargombak). A fonal alaku bakteriumokat gyakran tok veszi korul, mely szamos egyedulallo sejtet is tartalmaz. Bizonyos fajok, mint a
Nocardia
nemzetseg
, osszetett elagazo fonalakat formaz, mely megjelenesre hasonlit a
gombak
miceliumara
.
[33]
 A
prokariota
sejtek merettartomanya, a tobbi organizmushoz es
biomolekulahoz
viszonyitva
A
prokariota
sejtek merettartomanya, a tobbi organizmushoz es
biomolekulahoz
viszonyitva
A bakteriumok gyakran tapadnak kulonfele feluletekhez es egybefugg? bevonatot,
biohartyat
(biofilm) vagy
bakteriumsz?nyeget
alkotnak. A bevonat vastagsaga nehany mikrometert?l a fel meterig terjedhet, es benne tobb bakteriumfaj, valamint a
Protista
es az
Archeak
csoport kepvisel?i is el?fordulhatnak. A bevonatban el? bakteriumok sejtjei es a sejten kivuli komponensek bonyolult modon rendez?dnek el, masodlagos strukturakat, peldaul mikrokoloniakat hoznak letre, melyeken keresztul csatornak rendszere biztositja, hogy a tapanyagok megfelel? modon jussanak el az egyes sejtekhez.
[34]
[35]
Termeszetes korulmenyek kozott, mint peldaul a foldben es a novenyek feluleten a bakteriumok tobbsege bevonatot alkotva talalhato meg.
[36]
A biohartya fontos a kronikus bakterialis fert?zeseknel, vagy a beultetett orvosi eszkozoknel fellep? fert?zeseknel, mert a bakteriumokat megvedi, es igy sokkal nehezebben pusztithatok el, mint az egyedi sejtek.
[37]
Neha meg osszetettebb morfologiai valtozasok is lehetsegesek. Aminosavhiany eseten a
myxobakterium
(Myxobacteria)
fajok sejtjei egymas fele vandorolnak, osszetapadnak es akar 500 mikrometer hosszu, fajra jellemz? alaku es szin?
term?testet
formaznak, melyekben kozel 100 000 bakteriumsejt talalhato.
[38]
A term?testben a bakteriumok mar kulon feladatokat is vegeznek a
tobbsejt? szervez?des
egyik egyszer? tipusakent. Peldaul kb. minden tizedik sejt a term?test felszinere vandorol, es egy specialis allapotu sejtte, un. myxosporava alakul. A myxosporak a kiszaradasnak es a karos kornyezeti felteteleknek jobban ellenallnak, mint a normalis sejtek (kitarto keplet).
[39]
 Tipikus bakteriumsejt felepitese
Tipikus bakteriumsejt felepitese
Sejten beluli strukturak
[
szerkesztes
]
A bakterialis sejtet
lipidmembran
, mas neven
sejtmembran
burkolja, mely egyreszt hatarolja a sejttartalmat, masreszt akadalyt kepez, es a tapanyagokat, feherjeket es a
citoplazma
egyeb eletfontossagu alkotoreszeit a sejten belul tartja. A sejtmembran szoros kapcsolatban all a sejtet kivulr?l hatarolo
sejtfallal
. A foszfolipidekb?l es
feherjekb?l
allo kett?s hartya szerepe sokret?: a DNS a
mezoszomahoz
tapad a membranon; a legzesi
enzimek
is a membran lemezes beturemkedeseiben helyezkednek el, illetve a bioszintetikus, metabolikus reakciok egy resze is a hartya menten folyik.
Mivel prokariota szervezetek, nincsenek membrannal boritott
sejtszervecskek
(sejtorganellumok) a citoplazmaban, es igy keves sejten beluli strukturat tartalmaznak. Mindegyikukb?l hianyzik a
sejtmag
, a
mitokondrium
, a
szintest
es az eukariota sejtekben megtalalhato tobbi sejtszervecske, mint peldaul a
Golgi-keszulek
vagy az
endoplazmatikus retikulum
.
[40]
A bakteriumok nem rendelkeznek membrannal boritott
sejtmaggal
, orokit?anyaguk, a
DNS
rendszerint egy darab korkoros
kromoszoma
. Ez a citoplazmaban lev? szabalytalan formaju kepletben, az un.
nukleoidban
talalhato,
[41]
a hozzatapadt hisztonszer? feherjekkel es az
RNS
-sel egyutt. A 80%-os viztartalommal rendelkez? sejtplazmaban talalhatok (mint minden el? organizmus eseteben) a feherjeszintezist vegz?
riboszomak
, de ezek felepitese egyreszt elter az eukariotak es az archeak riboszomainak felepiteset?l,
[42]
masreszt szamuk joval nagyobb, mint az eukariotakban. A
Planctomycetes
rend tagjai kivetelesek abbol a szempontbol, hogy esetukben a nukleoidot membran veszi korbe, es rendelkeznek egyeb membrannal boritott sejtstrukturakkal is.
[43]
A bakteriumok egy resze sejten beluli tapanyag-raktarozo gomboket
(granulumokat)
kepez, melyek
glikogent
,
[44]
polifoszfatot
,
[45]
esetleg
kent
[46]
tartalmaznak. Ezek a granulumok lehet?ve teszik, hogy a bakteriumok ezeket az anyagokat kes?bbi hasznalatra elraktarozzak. Bizonyos bakteriumfajok, mint peldaul a fotoszintetizalo
cianobakterium
-fajok gazvezikulumokat kepeznek a sejten belul, melyekkel a sejtjeik felhajtoerejet szabalyozzak annak erdekeben, hogy optimalis feny- es tapanyagviszonyok koze keruljenek.
[47]
Sejten kivuli strukturak
[
szerkesztes
]
A sejtmembranon kivul helyezkedik el a bakterialis
sejtfal
, mely a bakteriumot vedi a kornyezeti hatasoktol es esetleg a gazdaszervezet
immunrendszere
ellen. A sejtfal emellett fontos szerepet jatszik a sejt magas
ozmozisnyomasanak
fenntartasaban, ami akar a legkori
nyomas
tizenotszorose is lehet. A sejtfal f? alkotoresze
peptidoglikan
, azaz olyan molekulak, amelyekben a peptidekhez poliszacharidlancok kapcsolodnak
kovalens kotessel
.
[48]
A bakterialis peptidoglikan (mas neven murein) terhalos szerkezet?:
poliszacharidlancai
D-aminosavakat
tartalmazo peptidekkel van keresztulkotve.
[49]
A bakterialis sejtfal elter a
novenyek
es a
gombak
sejtfalatol, mivel azok
cellulozbol
, illetve
kitinb?l
allnak.
[50]
A bakteriumok sejtfala az archeak sejtfalatol is kulonbozik, mivel azok sejtfala nem tartalmaz peptidoglikanokat. A sejtfal alapvet? fontossaggal bir a tuleles szempontjabol: a
penicillinszarmazekok
eppen azaltal teszik lehet?ve a bakteriumok elpusztitasat, hogy gatoljak a peptidoglikan szinteziset.
[50]
Leegyszer?sitve ket kulonboz? tipusu sejtfal talalhato a bakteriumokban, ezek alapjan
Gram-pozitiv
es
Gram-negativ
bakteriumokra lehet felosztani a fajokat. A nev a bakteriumfajok osztalyozasara regota hasznalatos
Gram-festes
eredmenyere utal.
[51]
A Gram-pozitiv bakteriumok sejtfala vastag, sok peptidoglikan- es lipoteichnoinsav-reteget tartalmaz. A Gram-negativ bakteriumok ezzel szemben viszonylag vekony sejtfallal rendelkeznek, mely csak nehany reteg peptidoglikanbol all, melyet
lipopoliszacharidokat
es
lipoproteineket
tartalmazo masodik lipidmembran burkol. A legtobb bakterium a Gram-negativ csoportba tartozik, csak a
Firmicutes
es
Actinobacteria
torzs tagjainak van Gram-pozitiv sejtfala.
[52]
A felepitesbeli kulonbsegek elter? erzekenyseget eredmenyeznek az antibiotikumokkal szemben, peldaul a
vankomicin
csak Gram-pozitiv bakteriumokat tud elpusztitani, es Gram-negativ patogenekkel, mint peldaul a
Haemophilus influenzae
vagy a
Pseudomonas aeruginosa
fajokkal szemben hatastalan.
[53]
Szamos bakterium eseteben egy merev szerkezet? feherjemolekulakbol allo S-reteg boritja a sejtet.
[54]
Ez a reteg kemiai es fizikai vedelmet biztosit a sejtfelszinnek, es egyben a
makromolekulak
diffuziojat
akadalyozza. Az S-retegnek mas, meg kevesse ismert funkcioi is vannak. Ismeretes peldaul, hogy a
Campylobacter
fert?z?kepessegehez hozzajarul, es a
Bacillus stearothermophilus
eseteben felszini enzimeket is tartalmaz.
[55]
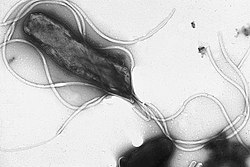 Helicobacter pylori
elektronmikroszkopos felvetele, jol lathatoak a sejtfelszini ostorok.
Helicobacter pylori
elektronmikroszkopos felvetele, jol lathatoak a sejtfelszini ostorok.
Az
ostorok
kb. 20
nanometer
atmer?j?, es akar 20 mikrometer hosszusagu merev feherjekepz?dmenyek, melyek az aktiv helyvaltoztatast szolgaljak. A mozgashoz szukseges energiat az elektrokemiai gradienst kovetve a sejtmembranon athalado ionok szolgaltatjak.
[56]
A
csillok
2?10 nanometer atmer?j? es legfeljebb nehany mikrometer hosszu feherjefonalak. A sejtfelszint beborito csillok finom sz?rzetre emlekeztetnek az elektronmikroszkopban. Mai ismereteink alapjan a szilard feluletekhez vagy mas sejtekhez torten? tapadasban jatszanak szerepet, es egyes patogen bakteriumok fert?z?kepesseget is meghatarozzak.
[57]
A pilusok az ostoroknal nemileg nagyobb sejtfuggelekek, melyeken keresztul az osszetapadt bakteriumsejtek genetikai anyagot cserelnek egymassal (
konjugacio
, l. kes?bb).
[58]
A bakteriumok egy reszet korulvev? tokok vagy nyalkaburkok szerkezetileg er?sen elter?ek: megtalalhato kozottuk a sejten kivuli strukturalatlan
polimert?l
kezdve a szigoruan strukturalt tokig vagy
glikokalix
burokig minden. Ezek a strukturak megvedhetik a sejteket mas sejtek, peldaul
makrofagok
altal torten? bekebelezest?l.
[59]
Antigenkent
szerepet jatszhatnak abban, hogy az immunrendszer rajtuk keresztul felismeri a betolakodokat, de segitik a kulonboz? feluletekhez torten? tapadast es a biohartyak kepzeset is.
[60]
Ezeknek a sejten kivuli strukturaknak az osszeallitasa a bakterialis kivalasztorendszerekt?l fugg. Ezek a rendszerek juttatjak ki a feherjeket a citoplazmabol a
periplazmaba
vagy a sejt kornyezetebe. Szamos ilyen rendszer ismert, es mivel a patogenek fert?z?kepessegenek szempontjabol meghatarozoak, intenziven kutatjak ezeket.
[61]
 Gerincvel?i folyadekban noveked?
Bacillus anthracis
(biborszin? festes)
Gerincvel?i folyadekban noveked?
Bacillus anthracis
(biborszin? festes)
A Gram-pozitiv bakteriumok bizonyos nemzetsegei, mint peldaul a
Bacillusok
,
Clostridiumok
, peldaul
Clostridium difficile
,
Sporohalobacterek
,
Anaerobacterek
vagy a
Heliobacteriumok
sejtjei nyugvo allapotu kepletekke, un.
endosporakka
alakulhatnak.
[62]
Legtobbszor csak egyetlen endospora kepz?dik a bakteriumsejtben, amely az eredeti sejt pusztulasaval kerul a szabadba, igy celja nem a szaporodas, hanem a kedvez?tlen korulmenyek atveszelese (kitarto keplet). Az
Anaerobacter
fajok kepesek akar 7 endosporat kepezni egyetlen sejtben (kitarto es szaporito keplet).
[63]
Az endosporak kozepen talalhato a
citoplazma
a DNS-sel es a riboszomakkal, ezeket veszi korbe egy kuls? reteg (kereg), melyet egy at nem ereszt? merev burok zar korbe.
Az endosporaknak nincs anyagcsereje. Szels?seges fizikai es kemiai korulmenyeket kepesek atveszelni, peldaul er?s UV- vagy gamma-sugarzast, oldoszereket,
fert?tlenit?szereket
, h?seget, nyomast es kiszaradast.
[64]
S?t evmilliokig eletkepesek maradhatnak a nyugvo allapotban.
[65]
[66]
Az endosporaknak koszonhet?en a bakterium meg az ?rben talalhato vakuumot es sugarzast is tulelheti.
[67]
Az endosporakat kepez? bakteriumok kozott korokozok is akadnak: peldaul a
lepfene
elkaphato a
Bacillus anthracis
endosporainak belelegzesevel, vagy a mely sebbe jutott
Clostridium tetani
endospora
tetanuszt
okoz.
[68]
A magasabb rend? organizmusokkal szemben a bakteriumok
anyagcsereje
nagyon valtozatos kepet mutat.
[69]
Hagyomanyosan az anyagcsere jellegzetessegei alapjan hataroztak meg a
rendszertanukat
, de ez az osztalyozas gyakran elter a modern
genetikai
osztalyozastol.
[70]
A bakterialis anyagcsere durva felosztasanak alapjat az adja, hogy az adott bakterium a novekedeshez milyen szen- es energiaforrast hasznal, valamint az energiatermel? folyamatok soran mely anyagok es vegyuletek adjak az elektront (elektrondonor) es mely anyagok es vegyuletek kapjak a vegen az elektront (elektronakceptor).
[71]
Szenforras szempontjabol a bakteriumok lehetnek
heterotrofok
, azaz a kornyezetukben talalhato szerves szenvegyuleteket hasznaljak szen- es energiaforraskent, vagy
autotrofok
, azaz szenforraskent a kornyezet szen-dioxidjat hasznaljak. Az autotrof bakteriumok tipikus kepvisel?i a fotoszintetizalo
cianobakteriumok
, zold kenbakteriumok es reszben a biborbakteriumok, de autotrof sok kemolitotrof faj is, mint peldaul a nitrifikalo es a kenoxidalo bakteriumok.
[72]
Energiaforras szempontjabol a bakteriumok vagy fotoszintetizalok, azaz
fotoszintezis
utjan a fenyb?l nyerik az energiat, vagy
kemoszintetizalok
, azaz kemiai vegyuletekb?l nyerik az energiat. A kemoszintetizalokat tovabb szokas bontani
kemolitotrofokra
(a legzeshez szervetlen elektrondonort hasznalnak) es
kemoorganotrofokra
(a legzeshez szerves elektrondonort hasznalnak). Kemolitotrof bakteriumok eseteben a leggyakoribb energiaforras a
hidrogen
,
szen-monoxid
,
ammonia
(ennek eredmenye a
nitrifikalas
), esetleg vasion, vagy mas redukalt femion, es szamos
kenvegyulet
. A legtobb kemolitotrof szervezet autotrof, mig a kemoorganotrof szervezetek heterotrofok.
Elektrondonorok es -akceptorok tekinteteben: a kemiai vegyuletek energiaforraskent torten? felhasznalasa soran az
oxidalodo
anyagbol az elektronok a vegs? elektronfelvev?nek kerulnek atadasra,
redukcios
folyamat soran. Ebben a reakcioban energia szabadul fel, mely az anyagcsere soran felhasznalhato. Az
aerob
el?lenyek eseteben az
oxigen
az elektronfelvev?.
Anaerob
el?lenyek eseteben mas szervetlen vegyulet, peldaul
nitrat
,
szulfat
, vagy
szen-dioxid
az elektronfelvev?, aminek eredmenye az okologiai szempontbol is fontos
denitrifikalas
, kentelenites es acetogenezis
[73]
[74]
). Leteznek
fakultativ anaerob
bakteriumok, melyek ha nem all rendelkezesre vegs? elektronfelvev?,
erjedessel
biztositjak eletm?kodesuket. Ennek soran cukrokbol, vagy egyeb magas energiatartalmu vegyuletekb?l allitanak el? az erjedes tipusatol fugg?en
tejsavat
,
etil-alkoholt
,
hidrogent
,
vajsavat
vagy egyeb vegtermekeket.
A kornyezetszennyezesre adott biologiai valaszban is fontosak ezek a folyamatok, peldaul szulfatredukalo bakteriumok termelik a kornyezetben talalhato kulonosen mergez?
higanyvegyuletek
(metil-, es
dimetil-higany
) nagy reszet.
[75]
Az aerob fotoszintetizalo es a kemolitotrof szervezetek eseteben oxigen az elektronfelvev?, de anaerob korulmenyek kozott az oxigen helyett szervetlen vegyuleteket hasznalnak.
Kulonleges eset a metanotrof bakteriumok esete, amikor a
metangaz
szolgaltatja az elektronokat es egyben szenforras is.
[76]
A fotoszintezis soran megkotott szen-dioxid mellett nehany bakterium a legkori
nitrogent
koti meg a
nitrogenaz
enzimmel (ilyenek peldaul a talajban el?
nitrifikalo bakteriumok
). A nitrogenkot? kepesseg csaknem mindegyik fent felsorolt anyagcseretipussal parosulhat.
[77]
A tobbsejt? szervezetekt?l elter?en a bakteriumsejtek meretenek novekedese es
osztodasa
szorosan osszefugg egymassal. A bakteriumok egy bizonyos meretig novekednek, majd ketteosztodnak.
[78]
A folyamat
ivartalan szaporodas
, amit a bakteriumok eseteben
hasadasnak
szokas nevezni, megkulonboztetve a valodi sejtmaggal es kromoszomakkal rendelkez? eukariota sejtek
sejtosztodasatol
.
[79]
Optimalis korulmenyek eseten a bakteriumok rendkivul gyorsan novekednek es osztodnak, akar 9,8 perc alatt is megduplazodhat egy bakteriumpopulacio.
[80]
A sejtosztodas soran ket azonos utodsejt keletkezik. Nehany ivartalanul szaporodo bakterium ennel bonyolultabb kepleteket alakit ki a szaporodas soran, ezek az ujonnan letrejott utodsejtek eloszlasat szabalyozzak. Erre jo pelda a
myxobakteriumok
term?teste, a
Streptomyces
fajok
hifai
vagy a
bimbozas
, mely soran egy kituremked? resz letorik, es igy jon letre az utodsejt.
 Bakteriumtenyeszet
agaragar-taptalajon
egy
Petri-cseszeben
Bakteriumtenyeszet
agaragar-taptalajon
egy
Petri-cseszeben
Laboratoriumban a bakteriumokat rendszerint szilard vagy folyekony kozegben tenyesztik. Tiszta tenyeszetek izolalasahoz szilard kozeget, peldaul
agaragar-taptalajt
, a szaporodas meresehez vagy nagy mennyiseg? sejt el?allitasahoz folyekony kozeget hasznalnak. A folyekony kozeget folyamatosan keverik, hogy egyenletes
sejtszuszpenziot
kapjanak, amit konny? tovabb szaporitani es szallitani, viszont nehez bel?le egy-egy bakteriumcsoportot elkuloniteni. A bakteriumok azonositasa tortenhet szelektiv (peldaul bizonyos tapanyagok vagy antibiotikumok hozzaadasaval vagy kihagyasaval el?allitott) kozeg felhasznalasaval.
[81]
Nagy mennyiseg? bakterium gyors es olcso el?allitasahoz a legtobb laboratoriumi technika b?segesen adagolja a tapanyagokat. Termeszetes korulmenyek kozott azonban a tapanyagok mennyisege veges, ami azt is jelenti, hogy a bakteriumok nem tudnak korlatlanul szaporodni. A tapanyagok korlatossaga kulonboz? novekedesi strategiakhoz vezetett (
r-K
strategia). Nehany organizmus rendkivul gyors szaporodasra kepes, ha a tapanyagok rendelkezesre allnak (
r
-strategia). Erre jo pelda az algaviragzas jelensege, amely a nyari melegben oxigenszegennye valt, de tapanyagokban gazdag sekely tavakban katasztrofalis mereteket is olthet a
cianobakteriumok
(regi nevukon kekmoszatok) elszaporodasaval.
[82]
Mas bakteriumok inkabb utodaik tulelesi eselyet novelik (
K
-strategia). Peldaul a
Streptomyces
fajok kulonfele
antibiotikumokat
termelnek, amivel mas mikroorganizmusok novekedeset gatoljak.
[83]
A termeszetben sokan valasztjak a kozossegi eletet (peldaul
biohartya
), amely segithet a taplalkozasban es vedelmet is nyujthat,
[36]
de letfeltetel is lehet (peldaul ilyen a
szintrofia
jelensege, amikor ket mikroorganizmus kolcsonosen fugg egymas anyagcseretermeket?l).
[84]
A bakteriumpopulaciok novekedese harom f? szakaszra oszthato. Amikor bakteriumok kerulnek a megfelel? tapanyaggal ellatott kornyezetbe, a sejteknek el?szor alkalmazkodniuk kell az uj kornyezethez. A novekedes els? szakasza a lappango fazis, a lassu novekedes szakasza, mikor a sejtek felkeszulnek es atallnak a gyors novekedesre a megfelel? enzimrendszerek, transzportfeherjek szintetizalasaval.
[85]
A masodik novekedesi szakasz a logaritmikus fazis, mas neven exponencialis fazis. Ennek jellemz?je a gyors, exponencialis novekedes. Az egyedszam id?egyseg alatti novekedeset mutatja a
novekedesi rata,
az egyedszam megduplazodasat pedig a
generacios id?.
Ebben a fazisban a sejtek a tapanyagokat maximalis sebesseggel hasznaljak fel az anyagcserejukben, a gyors reprodukcio miatt a
genetikai allomany
megkett?z?dese folyamatosan zajlik. Meg miel?tt az els? kett?z?des vegbemenne, megkezd?dik a kovetkez?. Ezert egy id?ben tobb replikacios villat is lathatunk a
DNS
-en. Ez egeszen addig tart, mig a tapanyagok el nem kezdenek fogyni, korlatozva a szaporodast. Az utolso fazis a stacioner vagy vesztegl? fazis, melyet a tapanyaghiany okoz. A sejtek csokkentik az anyagcserejuket, es lebontjak a nem eletfontossagu sejtfeherjeket. A stacioner fazis a gyors novekedes allapotabol a stresszre adott valaszallapotba torten? atmenet, melynek soran megnovekedik a DNS-javitassal, az
antioxidans
-anyagcserevel es a tapanyagszallitassal osszefugg? genek aktivitasa.
[86]
A bakteriumok tobbsegenek egyetlen, korkoros
kromoszomaja
van. Meretet tekintve a
Mycoplasma genitalium
korokozo 580 ezer
bazisparral
a legkisebb,
[87]
mig 12,2 millio bazisparral a talajlako
Sorangium cellulosum
a legnagyobb
[88]
ismert bakterialis kromoszoma. A
spirochaetak
(peldaul a
Lyme-kor
korokozoja, a
Borrelia burgdorferi
) ett?l elter?en linearis kromoszomaval rendelkeznek.
[89]
A bakterialis kromoszoma
hisztonok
helyett hisztonszer? feherjeket tartalmaz. A bakteriumsejtben el?fordulhatnak plazmidok is, olyan kis meret?, kor alaku orokl?d?
DNS-darabok
, amelyek nem reszei a kromoszomanak. A plazmidok
antibiotikum-rezisztenciaert
, fert?z?kepessegert felel?s
geneket
is hordozhatnak. A bakterialis DNS egy resze viruseredet?. Szamos bakterialis virus, azaz
bakteriofag
ismeretes. Nehany egyszer?en megfert?zi es elpusztitja a bakteriumokat, masok beepulnek a bakterialis kromoszomaba. A bakteriofag tartalmazhat olyan geneket, melyek a gazda
fenotipusat
is befolyasoljak. Peldaul az
Escherichia coli
O157:H7
es a
Clostridium botulinum
evolucioja soran bakteriofag
toxingenek
valtoztattak at az eredetileg artalmatlan bakteriumokat halalos korokozokka.
[90]
[91]
A bakteriumok ivartalanul szaporodnak, igy utodaik genetikai allomanya megegyezik. A bakteriumok evolucioja a genetikai anyagban bekovetkezett
rekombinacio
es
mutacio
reven el?allo modosulasok szelekciojaval valosul meg. Mutacio a DNS hibas masolasakor, vagy
mutagenekkel
torten? erintkezeskor kovetkezik be. A bakteriumfajok, s?t az egy fajba tartozo torzsek mutacios rataja is nagyon elter? lehet.
[92]
Mutaciohoz vezethet a stressz is: ilyenkor bizonyos, a novekedest korlatozo folyamatokkal osszefuggesben allo geneknek novekedik meg a mutacios rataja.
[93]
Sok bakteriumnal megfigyeltek az orokit?anyag sejtek kozotti atvitelet (horizontalis gentranszfer). Ennek harom f? modja van. A
transzformacio
soran a bakterium kepes a kornyezeteben lev? DNS-t felvenni. Az igy felvett DNS gyakran nem kerul be a bakterium kromoszomajaba, hanem
plazmidkent
talalhato meg a sejtben. Genek kerulhetnek be a bakteriumba a
transzdukcio
utjan is, ekkor egy bakteriofag illeszt a bakterialis kromoszomaba idegen DNS-t. A harmadik mod a
konjugacio
, amikor kozvetlen sejtkapcsolat utjan cserel?dik ki DNS. A horizontalis gentranszfer termeszetes korulmenyek kozott gyakori jelenseg.
[94]
A genatvitelnek jelent?s szerepe van az
antibiotikum-rezisztencia
szempontjabol is, mivel lehet?ve teszi a rezisztenciaert felel?s genek gyors atadasat akar kulonboz? korokozo fajok kozott is.
[95]
 Bakteriumostorok elrendez?desei: A. monotrich; B. lofotrich; C. amfitrich; D. peritrich.
Bakteriumostorok elrendez?desei: A. monotrich; B. lofotrich; C. amfitrich; D. peritrich.
A bakteriumok
ostorral
, csuszassal, rangatozo mozgassal, vagy a felhajtoer? valtoztatasaval kepesek helyuket megvaltoztatni.
[96]
A bakteriumok kozott egyedulallo modon a
spirochaetaknak
ostorhoz hasonlo kepletei, un. axialis filamentumai vannak, melyek nem a sejthartyaban, hanem a sejthartya es a kuls? membran kozotti periplazmatikus terben talalhatok. Jellegzetes spiralisan csavart testuk van, mely mozgas kozben meghajlik.
[96]
Az ostorok szama es a sejt felszinen torten? elrendez?dese elter a bakteriumfajoknal. Vannak, melyeknek egyetlen ostora van (
monotrich
), leteznek fajok, melyeknel a sejt ket vegen van egy-egy ostor (
amfitrich
), esetleg a sejt egyik vegen egy halomban van sok ostor (
lofotrich
), vagy a sejt mindket vegen tobb flagellum talalhato (
amfilofotrich
), es ismertek fajok, melyeknel a sejt teljes feluletet beboritjak az ostorok (
peritrich
).
 A bakterialis ostor felepitese
A bakterialis ostor felepitese
Az el?lenyek mozgasat szolgalo valamennyi struktura kozul a bakteriumok ostoranak a szerkezetet es m?kodeset ismerjuk a leginkabb. Az ostor mintegy 20 feherjeb?l epul fel, es korulbelul masik 30 feherje jatszik szerepet a szabalyozasaban es elkeszuleseben.
[96]
Az ostort az ostor toveben elhelyezked? motor forgatja, amely a hajocsavarhoz hasonloan hajtja el?re a sejtet. Szamos bakterium (peldaul az
E. coli
) ket kulonboz? modon tud mozogni: el?rehalado mozgassal (uszas) es bukfencezessel. A bukfencezessel tudnak uj iranyba allni, es terben mozogni.
[97]
A mozgasra kepes bakteriumokat bizonyos ingerek vonzzak vagy taszitjak, ezt a viselkedest
taxis
utotaggal jelolik: peldaul
kemotaxis
,
fototaxis
vagy
magnetotaxis
.
[98]
[99]
A
myxobakteriumoknal
figyelhet? meg az a jelenseg, hogy az egyes bakteriumsejtek egyutt mozognak, mikozben a sejtekb?l hullamok formalodnak, melyekb?l kes?bb az endosporakat tartalmazo term?testek lesznek.
[39]
Ezek a bakteriumok csak akkor mozognak, ha szilard felszinen vannak, de peldaul az
E. coli
akar folyekony kozegben, akar szilard felszinen is kepes mozogni.
Nehany
Listeria
es
Shigella
faj a gazdasejten belul a gazdasejt
citoszkeletonjanak
segitsegevel mozog (amit egyebkent a sejt a sejtszervecskek mozgatasara hasznal). A sejtjeik egyik oldalanal el?segitik az
aktin
polimerizaciojat
, es a novekv? aktin filamentumok nyomjak a masik iranyba a bakteriumsejteket a gazdasejten belul.
[100]
Osztalyozasuk es azonositasuk
[
szerkesztes
]
Az
osztalyozas
celja, hogy leirja az egyes bakteriumfajok kozotti eltereseket az el?lenyek hasonlosagan alapulo csoportositassal es elnevezessel. A bakteriumok a sejtstrukturaik, anyagcserejuk vagy az olyan sejtalkotokban lev? kulonbsegek alapjan osztalyozhatoak, mint a DNS,
zsirsavak
, pigmentek, vagy az
antigenek
.
[81]
Bar ezek alapjan lehet?ve valt a bakteriumtorzsek azonositasa es osztalyozasa, nem volt vilagos, vajon a kulonbsegek fajok kozotti elteresek, vagy pedig egy fajon beluli elteresek. Ennek a bizonytalansagnak az az oka, hogy a legtobb bakteriumfajban nincsenek jol megfigyelhet? jellegzetes kepletek, valamint hogy az egymastol fuggetlen fajok kozott is letezik a horizontalis gentranszfer jelensege.
[107]
A horizontalis gentranszfer miatt a kozeli rokonsagban allo bakteriumoknak is egeszen elter? morfologiaja vagy anyagcsereje lehet. Annak erdekeben, hogy ez a bizonytalansag csokkenthet? legyen, a modern bakteriologia egyre inkabb a molekularis rendszertanra tamaszkodik, olyan technikakat felhasznalva, mint a
guanin
-
citozin
arany meghatarozasa, vagy olyan genek
DNS-szekvencialasa
, melyeket nem erintett komolyan a horizontalis gentranszfer (peldaul
rRNS
-genek).
[108]
 Gram-festes alkalmazasa
Streptococcus mutans
kimutatasara
Gram-festes alkalmazasa
Streptococcus mutans
kimutatasara
 Ziehl?Neelsen festes alkalmazasa
Mycobacterium tuberculosis
kimutatasara.
Ziehl?Neelsen festes alkalmazasa
Mycobacterium tuberculosis
kimutatasara.
A bakteriumok azonositasa kulonosen az orvostudomanyban jut nagy szerephez, ahol a megfelel? kezeles a fert?zest okozo bakteriumfaj ismeretet?l fugg. Emiatt a bakteriumok azonositasara szolgalo technikak fejl?deset dont?en befolyasolta az emberi korokozok azonositasanak surget? igenye.
A bakteriumokat kulonboz? festesi eljarasokkal szembeni viselkedes alapjan is szoktak csoportositani. Az egyik ilyen eljaras a
Gram-festes
, melyet
1884
-ben
Hans Christian Gram
fejlesztett ki. Ez a modszer a bakteriumokat a sejtfal strukturalis sajatossagai alapjan kuloniti el.
[51]
A festes soran
kristalyibolya
(vagy gencianaibolya)-festekkel festik meg a bakteriumkeszitmenyt, majd
etanollal
mosasi probat vegeznek. Gram-pozitiv bakteriumok eseten a festek nem moshato ki a sejtb?l, mig a Gram-negativ bakteriumoknal igen. A Gram-negativ bakteriumok lathatova tetele erdekeben tovabbi
fukszinos
festest alkalmaznak. A Gram-pozitiv bakteriumok lilanak latszanak, mig a Gram-negativ fajok rozsaszin?ek lesznek a festes utan. A Gram-festest a morfologiaval kombinalva a legtobb bakterium besorolhato 4 csoport egyikebe (Gram-pozitiv coccusok, Gram-pozitiv bacilusok, Gram-negativ coccusok, Gram-negativ bacilusok). Gram-pozitiv bakteriumok peldaul a tud?gyulladast okozo
Streptococcus pneumoniae
, Gram-negativ az emberi belcsatornaban el?
Escherichia coli
.
Bizonyos el?lenyek kimutatasara a Gram-festes helyett mas modszereket alkalmaznak, peldaul a
Mycobacterium tuberculosis
vagy a
Nocardia
egyedeinek kimutatasara elterjedt a
Ziehl?Nielsen-fele savallo festes
.
[109]
Ennel a modszernel forro
fenolos fukszinnal
festenek, ami ezekb?l a fajokbol nem oldodik ki sav hatasara
(savallo),
igy a sejtek a hatterhez kepest sotetebb szint vesznek fel. A
Corynebacterium dyphtheriae
foszfattartalmu volutinszemcseit, az un. Babe??Ernst-szemcseket lehet lathatova tenni a
Neisser-festessel
.
Szamos bakteriumot annak alapjan azonositanak, hogy milyen specialis kozegen tenyeszthet?ek. Ezeknek a technikaknak az az alapja, hogy bizonyos bakteriumfajok szaporodasat a kozeg el?segiti, mig mas fajok szaporodasat gatolja. Ezek a technikak gyakran bizonyos mintakra specifikusak, peldaul a kopetmintat ugy kezelik, hogy a
tud?gyulladast
okozo korokozot azonositsak, mig a szekletmintakat olyan szelektiv kozegeken tenyesztik, hogy ki tudjak mutatni a
hasmenes
korokozojat, egyuttal megakadalyozzak a nem korokozo bakteriumok szaporodasat. A normalis korulmenyek kozott steril mintakat (ver, vizelet, gerincvel?i folyadek) olyan kozegen tenyesztik, mely lehet?ve teszi az osszes bakterium szaporodasat.
[81]
[110]
Ezek mellett egyeb, peldaul
szerologiai
technikakat is bevetnek azonositas celjabol.
Mint a bakteriumok osztalyozasanal, az azonositasnal is novekv? szerephez jutnak a molekularis biologiai modszerek. Peldaul gyorsan novekszik a tenyeszteshez kepest gyors es pontos DNS alapu diagnosztikai modszer, a
polimeraz-lancreakcio
nepszer?sege.
[111]
Ezek a modszerek lehet?ve teszik azoknak az el?lenyeknek a felismereset es azonositasat is, amelyek ugyan anyagcserejuket tekintve aktivak, azonban nem osztodnak, es nem tenyeszthet?ek kulturaban.
[112]
Kapcsolat mas el?lenyekkel
[
szerkesztes
]
Nyilvanvalo egyszer?seguk dacara a bakteriumok bonyolult modon kepesek egyuttm?kodni mas el?lenyekkel. Ezeket a
szimbionta
kapcsolatokat feloszthatjuk
parazita
,
mutualista
es
kommenzalista
tipusuakra. A bakteriumok kis merete miatt a kommenzalizmus kozonseges, allatokon es novenyeken ugyanolyan jol novekednek, mint barmilyen mas feluleten.
Mutualista egyuttm?kodes
[
szerkesztes
]
A rizoszfera (a novenyi gyokerek altal befolyasolt talajresz)
nitrifikalo bakteriumai
legkori
nitrogent
kotnek meg,
[113]
igy konnyen felvehet? nitrogent biztositanak olyan novenyeknek, amelyek maguktol nem kepesek a molekularis
nitrogen
megkotesere.
Szamos mas bakterium el szimbiozisban az emberrel es mas el?lenyekkel. A normal emberi belfloraban jelen lev? korulbelul 1000-fele bakterium nagyban hozzajarul a bel immunrendszerehez,
folsav
-,
K-vitamin
, es
biotin
-szintezisehez, valamint a kulonben nem vagy nehezen emeszthet?
szenhidratok
emesztesehez.
[114]
[115]
Lactobacillus
fajok segitik a tejfeherjek tejsavva alakitasat a belben.
[116]
A belflora hasznos bakteriumai gatoljak a potencialisan korokozo bakteriumok szaporodasat (altalaban
kompetitiv kizaras
reven). Ezeket a hasznos bakteriumokat, mint
probiotikus
tapanyag-kiegeszit?ket forgalmazzak is.
[117]
 Treponema pallidum,
a szifilisz korokozoja
Treponema pallidum,
a szifilisz korokozoja
Korokozonak tekintjuk, ha a bakteriumok parazita egyuttm?kodest alakitanak ki mas el?lennyel. A
fert?zes
letrejohet erintkezessel, taplalekkal, leveg? vagy viz utjan.
A korokozo bakteriumoknak a
virusok
mellett nagy szerepuk van a
betegsegek
,
fert?zesek
kialakitasaban. Bakteriumok allnak szamos emberi megbetegedes mogott: kulonfele
gyulladasok
, mint a
mandulagyulladas
,
kozepfulgyulladas
,
sz?rtusz?gyulladas
,
pattanas
;
gyomorrontast
,
hasmenest
okozo fert?zesek, mint a
szalmonella
,
campylobacteriosis
,
verhas
,
kolera
; olyan ismert betegsegek, mint a
pestis
,
lepra
, TBC (
gum?kor
),
skarlat
,
difteria
,
tifusz
,
tetanusz
(vermergezes) es szamos
nemi betegseg
, mint a
gonorrhea
(tripper, kanko) vagy a
verbaj
(szifilisz). Bizonyos betegsegeknel sok evvel a betegseg leirasa utan derul ki, hogy korokozo all a hatterben, erre pelda, hogy sokaig nem volt egyertelm?, hogy a gyomorfekely hattereben a
Helicobacter pylori
bakterium all. A novenyi korokozokra pelda az almafelek
t?zelhalasos
megbetegedeseert felel?s
Erwinia amylovora
bakterium.
Mindegyik korokozo jellegzetes modon lep kapcsolatba a gazdaszervezettel. A
Staphylococcus
es
Streptococcus
peldaul b?rfert?zest, tud?gyulladast vagy akar
szepszist
okozhat.
[118]
Emellett ezek az el?lenyek a normalis emberi flora reszei, a b?ron es az orrban is megtalalhatoak es semmifele megbetegedest nem okoznak. Mas bakteriumok, peldaul a csak mas el?lenyek sejtjein belul noveked? es szaporodo
Rickettsia
fajok kivetel nelkul betegsegeket okoznak (a Rickettsiak egyik faja okozza a tifuszt). A sejten beluli parazitakhoz tartoznak a
Chlamydia
fajok, melyek a tud?gyulladasert vagy a hugyuti fert?zesekert felel?sek, es talan a szivkoszoruer megbetegedesekben is reszuk van.
[119]
Vegul leteznek az opportunista korokozok (mint a
Pseudomonas aeruginosa
,
Burkholderia cenocepacia
,
vagy a
Mycobacterium avium
), melyek csak akkor okoznak megbetegedest, ha a immunhianyos vagy cisztas fibrozisban szenved? embereket fert?znek meg.
[120]
[121]
A bakterialis fert?zesek kezelhet?ek
antibiotikumokkal
, melyeket
baktericidnek
nevezunk, ha elpusztitjak a bakteriumokat, vagy
bakteriosztatikusnak
hivunk, ha csak a szaporodasukat gatolja. Szamos kulonboz? tipusu antibiotikum letezik, mindegyikuk valamilyen modon gatol egy, a korokozoban es a gazdaszervezetben elter?en m?kod? folyamatot. Erre pelda a
chloramphenicol
es a
puromycin
, melyek a bakterialis riboszomak m?kodeset gatoljak, de nincsenek hatassal az ett?l elter? eukariota riboszomara.
[122]
Az antibiotikumokat az emberi megbetegedesek kezelese mellett az intenziv allattenyesztes soran is hasznaljak, annak erdekeben, hogy serkentsek az allatok novekedeset. Ezzel azonban hozzajarulhatnak a bakteriumpopulaciokban a gyors
antibiotikum-ellenallas
kialakulasahoz.
[123]
 191 bakterium
filogenetikai
eloszlasanak illusztralasa
191 bakterium
filogenetikai
eloszlasanak illusztralasa
- Higienia
?
Jarvany
idejen kerulni kell a tomegeket, gyakran kell tisztalkodni,
kezet mosni
. Csak ivovizb?l szabad inni, az elelmiszereket pedig lehet?leg frissen kell fogyasztani. Az orvosi m?szereket hasznalat el?tt
sterilizalni
kell. Ved?oltas, injekcio beadasa, sebeszeti beavatkozas el?tt a b?rt
alkohollal
kell fert?tleniteni.
- Jo kondicio ? Kevesbe betegszunk meg, ha kozerzetunk, testi, lelki allapotunk jo, ha eleget mozgunk, egeszsegesen taplalkozunk.
- Ved?oltas ? A bakteriumos megbetegedesek megel?zesenek is eszkoze lehet a
ved?oltas
.
- Antibiotikumok
? A mar kialakult bakteriumos megbetegedesek ellenszere, bakteriumol?, vagy szaporodasat gatlo anyag.
Figyelem
: Ez egy ketel? fegyver; csak akkor segit, ha az orvosi el?irast pontosan betartjuk, mert a gyogyszerkutatok a leghatasosabb gyogyszermennyiseg, beszedesi id?szakok es osszid?tartam osszesseget kiserletileg allapitottak meg ugy, hogy a kezeles a bakteriumkoncentraciot annyira csokkentse, hogy a maradekkal egyeni immunrendszerunk konnyen elbanhasson. Ha a beteg, magat jobban erezve a gyogyszer bevetelet elhanyagolja, vagy a kurat nem fejezi be, akkor a bakteriumok megmarado hanyada ismet felszaporulhat.
Darwin
termeszetes kivalasztodas
elmelete szerint az az egyed marad meg, amely az adott korulmenyekhez gyorsabban alkalmazkodik. Itt a megmarado bakteriumok jobban ellenalltak a kezelesnek, vagyis akaratunk ellenere a kezeles elhanyagolasaval egy antibiotikum-ellenallo csoportot fejlesztettunk ki.
Ugyeljunk tehat az orvos altal el?irt adagolas pontos betartasara, hogy ne segitsuk el? ellenallo bakteriumok kifejl?deset.
Jelent?seguk az iparban es a technologiaban
[
szerkesztes
]
Az emberiseg evezredek ota hasznalja a bakteriumokat (az
eleszt?kkel
es
peneszgombakkal
egyutt) olyan alapvet? elelmiszerek keszitesere, mint a
bor
,
sajtok
,
savanyusag
,
ecet
,
szojaszosz
, savanyu kaposzta vagy
joghurt
.
[124]
[125]
Figyelemremelto a bakteriumoknak azon kepessege, ahogy szerves anyagokat kepesek lebontani. Emiatt az iparban
hulladekfeldolgozasra
,
szennyviztisztitasra
hasznositjak ?ket, de specialis
szerves alapanyagu uveget
is kristalyosittatnak bakteriumokkal. A
k?olajban
lev?
szenhidrogeneket
lebonto bakteriumokat olajfoltok megszuntetesere hasznaljak.
[126]
Sikeresen alkalmaztak ezt a modszert az
1989
-es
Exxon Valdez
hajokatasztrofa olajszennyezesenel is, amikor m?tragyat szortak ki a Prince William obolben, hogy el?segitsek a termeszetesen is jelenlev? bakteriumok szaporodasat. Ipari szennyez?anyagok lebontasara is hasznalnak bakteriumokat.
[127]
A vegyiparban gyogyszerek es mez?gazdasagi termekek el?allitasaban szinten bakteriumok m?kodnek kozre.
[128]
A biologiai vedekezes soran a novenyved?szerek helyett is hasznalhatoak bakteriumok. Leggyakrabban a
Bacillus thuringiensis
nev? Gram-pozitiv talajlako bakterium valamelyik alfaja jut szerephez, mivel ezek a
Lepidoptera
rendre (lepkek, molyok) specifikus ?rovarirtok”.
[129]
Mivel az emberre, az el?vilagra es a hasznos rovarokra kicsi vagy semmilyen karos hatassal nincsenek, kornyezetbarat rovarirtonak tekinthet?ek.
[130]
[131]
A gyors novekedesuk es a konny? manipulalhatosaguk miatt a bakteriumok a
molekularis biologia
, a
genetika
es a
biokemia
?igaslovai”. A bakterialis DNS modositasaval, es az ennek eredmenyekent megvaltozo
fenotipus
tanulmanyozasaval a tudosok kepesek a
genek
,
enzimek
funkciojat es az anyagcsere utjait meghatarozni, es ezeket az eredmenyeket a magasabb rend? szervezetekre is alkalmazni.
[132]
A
biotechnologiaban
a bakterialis anyagcsere es genetika megertese teszi lehet?ve, hogy olyan modositott bakteriumokat allitsanak el?, melyek terapias celbol kepesek
inzulin
,
novekedesi faktorok
vagy
antitestek
el?allitasara.
[133]
[134]
- Pesti, Miklos.
Altalanos mikrobiologia
. Budapest?Pecs: Dialog Campus Kiado (2001).
ISBN 963-9123-71-4
- Gergely, Lajos.
Orvosi mikrobiologia
. Budapest: Semmelweis Kiado (1999).
ISBN 963-8154-93-4
- de Kruif, Paul.
Bacillusvadaszok
. Budapest: Konyvbaratok Szovetsege (1931)
- ↑
Bacteria (eubacteria)
.
Taxonomy Browser
. NCBI. (Hozzaferes: 2008. szeptember 10.)
- ↑
Fredrickson J, Zachara J, Balkwill D, et al (2004). ?
Geomicrobiology of high-level nuclear waste-contaminated vadose sediments at the hanford site, washington state
”.
Appl Environ Microbiol
70
(7), 4230?41. o. [2008. szeptember 29-i datummal az
eredetib?l
archivalva].
PMID 15240306
. (Hozzaferes: 2006. december 18.)
- ↑
Whitman W, Coleman D, Wiebe W (1998). ?
Prokaryotes: the unseen majority
”.
Proc Natl Acad Sci U S A
95
(12), 6578?83. o. [2008. marcius 5-i datummal az
eredetib?l
archivalva].
PMID 9618454
. (Hozzaferes: 2006. december 18.)
- ↑
Rappe M, Giovannoni S. ?The uncultured microbial majority”.
Annu Rev Microbiol
57
, 369?94. o.
PMID 14527284
.
- ↑
Sears C (2005). ?A dynamic partnership: Celebrating our gut flora”.
Anaerobe
11
(5), 247?51. o.
PMID 16701579
.
- ↑
Ishige T, Honda K, Shimizu S (2005). ?Whole organism biocatalysis”.
Curr Opin Chem Biol
9
(2), 174?80. o.
PMID 15811802
.
- ↑
Woese C, Kandler O, Wheelis M (1990). ?
Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya
”.
Proc Natl Acad Sci U S A
87
(12), 4576?9. o.
PMID 2112744
.
- ↑
Leeuwenhoek nevet illet?en az Anton, Antony es Antonie van Leeuwenhoek irasforma is ismeretes
- ↑
Leeuwenhoek A (1753). ?
Part of a Letter from Mr Antony van Leeuwenhoek, concerning the Worms in Sheeps Livers, Gnats, and Animalcula in the Excrements of Frogs
”.
Philosophical Transactions (1683?1775)
22
, 509?18. o. [2010. januar 5-i datummal az
eredetib?l
archivalva]. (Hozzaferes: 2007. augusztus 19.)
- ↑
Leeuwenhoek A (1753). ?
Part of a Letter from Mr Antony van Leeuwenhoek, F. R. S. concerning Green Weeds Growing in Water, and Some Animalcula Found about Them
”.
Philosophical Transactions (1683?1775)
23
, 1304?11. o. [2010. januar 18-i datummal az
eredetib?l
archivalva]. (Hozzaferes: 2007. augusztus 19.)
- ↑
Etymology of the word “bacteria”
Online Etymology dictionary.
- ↑
The Nobel Prize in Physiology or Medicine 1905
Nobelprize.org
- ↑
O'Brien S, Goedert J (1996). ?HIV causes AIDS: Koch's postulates fulfilled”.
Curr Opin Immunol
8
(5), 613?18. o.
PMID 8902385
.
- ↑
Thurston A (2000). ?Of blood, inflammation and gunshot wounds: the history of the control of sepsis”.
Aust N Z J Surg
70
(12), 855?61. o.
PMID 11167573
.
- ↑
Schwartz R (2004). ?Paul Ehrlich's magic bullets”.
N Engl J Med
350
(11), 1079?80. o.
PMID 15014180
.
- ↑
Biography of Paul Ehrlich
Nobelprize.org
- ↑
Woese C, Fox G (1977). ?Phylogenetic structure of the prokaryotic domain: the primary kingdoms”.
Proc Natl Acad Sci U S A
74
(11), 5088?90. o.
PMID 270744
.
- ↑
Woese C, Kandler O, Wheelis M (1990). ?
Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya
”.
Proc Natl Acad Sci U S A
87
(12), 4576?79. o.
PMID 2112744
.
- ↑
Schopf J (1994). ?
Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic
”.
Proc Natl Acad Sci U S A
91
(15), 6735?42. o.
PMID 8041691
.
- ↑
DeLong E, Pace N (2001). ?Environmental diversity of bacteria and archaea”.
Syst Biol
50
(4), 470?78. o.
PMID 12116647
.
- ↑
Brown J, Doolittle W (1997). ?
Archaea and the prokaryote-to-eukaryote transition
”.
Microbiol Mol Biol Rev
61
(4), 456?502. o.
PMID 9409149
.
- ↑
Di Giulio M (2003). ?
The universal ancestor and the ancestor of bacteria were hyperthermophiles
”.
J Mol Evol
57
(6), 721?30. o.
PMID 14745541
.
- ↑
Battistuzzi F, Feijao A, Hedges S. ?
A genomic timescale of prokaryote evolution: insights into the origin of methanogenesis, phototrophy, and the colonization of land.
”.
BMC Evol Biol
4
, 44. o.
PMID 15535883
.
- ↑
Poole A, Penny D (2007). ?Evaluating hypotheses for the origin of eukaryotes”.
Bioessays
29
(1), 74?84. o.
PMID 17187354
.
- ↑
Dyall S, Brown M, Johnson P (2004). ?Ancient invasions: from endosymbionts to organelles”.
Science
304
(5668), 253?7. o.
PMID 15073369
.
- ↑
McFadden G (1999). ?Endosymbiosis and evolution of the plant cell”.
Curr Opin Plant Biol
2
(6), 513?9. o.
PMID 10607659
.
- ↑
Lynn Sagan (1967). On the origin of mitosing cells.
J. Theoretical Biology
14
(3), 255?274.
PMID 11541392
- ↑
Schulz H, Jorgensen B. ?Big bacteria”.
Annu Rev Microbiol
55
, 105?37. o.
PMID 11544351
.
- ↑
Robertson J, Gomersall M, Gill P. (1975). ?
Mycoplasma hominis: growth, reproduction, and isolation of small viable cells
”.
J Bacteriol.
124
(2), 1007?18. o.
PMID 1102522
.
- ↑
Fritz I, Strompl C, Abraham W (2004). ?
Phylogenetic relationships of the genera Stella, Labrys and Angulomicrobium within the 'Alphaproteobacteria' and description of Angulomicrobium amanitiforme sp. nov
”.
Int J Syst Evol Microbiol
54
(Pt 3), 651?7. o. [2008. oktober 10-i datummal az
eredetib?l
archivalva].
PMID 15143003
. (Hozzaferes: 2006. december 27.)
- ↑
Cabeen M, Jacobs-Wagner C (2005). ?Bacterial cell shape”.
Nat Rev Microbiol
3
(8), 601?10. o.
PMID 16012516
.
- ↑
Young K (2006). ?
The selective value of bacterial shape
”.
Microbiol Mol Biol Rev
70
(3), 660?703. o.
PMID 16959965
.
- ↑
Douwes K, Schmalzbauer E, Linde H, Reisberger E, Fleischer K, Lehn N, Landthaler M, Vogt T (2003). ?Branched filaments no fungus, ovoid bodies no bacteria: Two unusual cases of mycetoma”.
J Am Acad Dermatol
49
(2 Suppl Case Reports), S170?3. o.
PMID 12894113
.
- ↑
Donlan R (2002). ?Biofilms: microbial life on surfaces”.
Emerg Infect Dis
8
(9), 881?90. o.
PMID 12194761
.
- ↑
Branda S, Vik S, Friedman L, Kolter R (2005). ?
Biofilms: the matrix revisited
”.
Trends Microbiol
13
(1), 20?26. o.
PMID 15639628
.
- ↑
a
b
Davey M, O'toole G (2000). ?
Microbial biofilms: from ecology to molecular genetics
”.
Microbiol Mol Biol Rev
64
(4), 847?67. o.
PMID 11104821
.
- ↑
Donlan RM, Costerton JW (2002). ?
Biofilms: survival mechanisms of clinically relevant microorganisms
”.
Clin Microbiol Rev
15
(2), 167?93. o.
PMID 11932229
.
- ↑
Shimkets L. ?Intercellular signaling during fruiting-body development of Myxococcus xanthus.”.
Annu Rev Microbiol
53
, 525?49. o.
PMID 10547700
.
- ↑
a
b
Kaiser D. ?Signaling in myxobacteria”.
Annu Rev Microbiol
58
, 75?98. o.
PMID 15487930
.
- ↑
Berg J., Tymoczko J. and Stryer L. (2002)
Biochemistry.
W. H. Freeman and Company
ISBN 0-7167-4955-6
- ↑
Thanbichler M, Wang S, Shapiro L (2005). ?The bacterial nucleoid: a highly organized and dynamic structure”.
J Cell Biochem
96
(3), 506?21. o.
PMID 15988757
.
- ↑
Poehlsgaard J, Douthwaite S (2005). ?The bacterial ribosome as a target for antibiotics”.
Nat Rev Microbiol
3
(11), 870?81. o.
PMID 16261170
.
- ↑
Fuerst J (2005). ?
Intracellular compartmentation in planctomycetes
”.
Annu Rev Microbiol
59
, 299?328. o.
PMID 15910279
.
- ↑
Yeo M, Chater K (2005). ?
The interplay of glycogen metabolism and differentiation provides an insight into the developmental biology of Streptomyces coelicolor
”.
Microbiology
151
(Pt 3), 855?61. o.
PMID 15758231
. (Hozzaferes: 2006. december 27.)
- ↑
Shiba T, Tsutsumi K, Ishige K, Noguchi T (2000). ?
Inorganic polyphosphate and polyphosphate kinase: their novel biological functions and applications
”.
Biochemistry (Mosc)
65
(3), 315?23. o.
PMID 10739474
.
- ↑
Brune DC. (1995). ?
Isolation and characterization of sulfur globule proteins from Chromatium vinosum and Thiocapsa roseopersicina
”.
Arch Microbiol
163
(6), 391?99. o.
PMID 7575095
.
- ↑
Walsby A (1994). ?
Gas vesicles
”.
Microbiol Rev
58
(1), 94?144. o.
PMID 8177173
.
- ↑
IUPAC-IUB Joint Commission on Biochemical Nomenclature (JCBN) (1986). ?Nomenclature of glycoproteins, glycopeptides and peptidoglycans”.
Eur. J. Biochem.
(159), 1?6. o.
- ↑
van Heijenoort J (2001). ?
Formation of the glycan chains in the synthesis of bacterial peptidoglycan
”.
Glycobiology
11
(3), 25R ? 36R. o.
PMID 11320055
.
- ↑
a
b
Koch A (2003). ?
Bacterial wall as target for attack: past, present, and future research
”.
Clin Microbiol Rev
16
(4), 673?87. o. [2008. szeptember 28-i datummal az
eredetib?l
archivalva].
PMID 14557293
. (Hozzaferes: 2006. december 27.)
- ↑
a
b
Gram, HC
(1884). ?Uber die isolierte Farbung der Schizomyceten in Schnitt- und Trockenpraparaten”.
Fortschr. Med.
2
, 185?189. o.
- ↑
Hugenholtz P (2002). ?
Exploring prokaryotic diversity in the genomic era
”.
Genome Biol
3
(2), REVIEWS0003. o.
PMID 11864374
.
- ↑
Walsh F, Amyes S (2004). ?Microbiology and drug resistance mechanisms of fully resistant pathogens.”.
Curr Opin Microbiol
7
(5), 439?44. o.
PMID 15451497
.
- ↑
Engelhardt H, Peters J (1998). ?Structural research on surface layers: a focus on stability, surface layer homology domains, and surface layer-cell wall interactions”.
J Struct Biol
124
(2?3), 276?302. o.
PMID 10049812
.
- ↑
Beveridge T, Pouwels P, Sara M, Kotiranta A, Lounatmaa K, Kari K, Kerosuo E, Haapasalo M, Egelseer E, Schocher I, Sleytr U, Morelli L, Callegari M, Nomellini J, Bingle W, Smit J, Leibovitz E, Lemaire M, Miras I, Salamitou S, Beguin P, Ohayon H, Gounon P, Matuschek M, Koval S (1997). ?Functions of S-layers”.
FEMS Microbiol Rev
20
(1?2), 99?149. o.
PMID 9276929
.
- ↑
Kojima S, Blair D. ?The bacterial flagellar motor: structure and function of a complex molecular machine”.
Int Rev Cytol
233
, 93?134. o.
PMID 15037363
.
- ↑
Beachey E (1981). ?
Bacterial adherence: adhesin-receptor interactions mediating the attachment of bacteria to mucosal surface
”.
J Infect Dis
143
(3), 325?45. o.
PMID 7014727
.
- ↑
Silverman P (1997). ?Towards a structural biology of bacterial conjugation”.
Mol Microbiol
23
(3), 423?9. o.
PMID 9044277
.
- ↑
Stokes R, Norris-Jones R, Brooks D, Beveridge T, Doxsee D, Thorson L (2004). ?
The glycan-rich outer layer of the cell wall of Mycobacterium tuberculosis acts as an antiphagocytic capsule limiting the association of the bacterium with macrophages
”.
Infect Immun
72
(10), 5676?86. o. [2008. szeptember 29-i datummal az
eredetib?l
archivalva].
PMID 15385466
. (Hozzaferes: 2006. december 27.)
- ↑
Daffe M, Etienne G (1999). ?The capsule of Mycobacterium tuberculosis and its implications for pathogenicity”.
Tuber Lung Dis
79
(3), 153?69. o.
PMID 10656114
.
- ↑
Finlay B, Falkow S (1997). ?
Common themes in microbial pathogenicity revisited
”.
Microbiol Mol Biol Rev
61
(2), 136?69. o.
PMID 9184008
.
- ↑
Nicholson W, Munakata N, Horneck G, Melosh H, Setlow P (2000). ?
Resistance of Bacillus endospores to extreme terrestrial and extraterrestrial environments
”.
Microbiol Mol Biol Rev
64
(3), 548?72. o.
PMID 10974126
.
- ↑
Siunov A, Nikitin D, Suzina N, Dmitriev V, Kuzmin N, Duda V. ?
Phylogenetic status of Anaerobacter polyendosporus, an anaerobic, polysporogenic bacterium
”.
Int J Syst Bacteriol
49 Pt 3
, 1119?24. o.
PMID 10425769
. (Hozzaferes: 2006. december 28.)
- ↑
Nicholson W, Fajardo-Cavazos P, Rebeil R, Slieman T, Riesenman P, Law J, Xue Y (2002). ?Bacterial endospores and their significance in stress resistance”.
Antonie Van Leeuwenhoek
81
(1?4), 27?32. o.
PMID 12448702
.
- ↑
Vreeland R, Rosenzweig W, Powers D (2000). ?Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal”.
Nature
407
(6806), 897?900. o.
PMID 11057666
.
- ↑
Cano R, Borucki M (1995). ?Revival and identification of bacterial spores in 25- to 40-million-year-old Dominican amber”.
Science
268
(5213), 1060?4. o.
PMID 7538699
.
- ↑
Nicholson W, Schuerger A, Setlow P (2005). ?The solar UV environment and bacterial spore UV resistance: considerations for Earth-to-Mars transport by natural processes and human spaceflight”.
Mutat Res
571
(1?2), 249?64. o.
PMID 15748651
.
- ↑
Hatheway C (1990). ?
Toxigenic clostridia
”.
Clin Microbiol Rev
3
(1), 66?98. o.
PMID 2404569
.
- ↑
Nealson K (1999). ?Post-Viking microbiology: new approaches, new data, new insights”.
Orig Life Evol Biosph
29
(1), 73?93. o.
PMID 11536899
.
- ↑
Xu J (2006). ?Microbial ecology in the age of genomics and metagenomics: concepts, tools, and recent advances”.
Mol Ecol
15
(7), 1713?31. o.
PMID 16689892
.
- ↑
Zillig W (1991). ?Comparative biochemistry of Archaea and Bacteria”.
Curr Opin Genet Dev
1
(4), 544?51. o.
PMID 1822288
.
- ↑
Hellingwerf K, Crielaard W, Hoff W, Matthijs H, Mur L, van Rotterdam B (1994). ?
Photobiology of bacteria
”.
Antonie Van Leeuwenhoek
65
(4), 331?47. o.
PMID 7832590
.
- ↑
Zumft W (1997). ?
Cell biology and molecular basis of denitrification
”.
Microbiol Mol Biol Rev
61
(4), 533?616. o.
PMID 9409151
.
- ↑
Drake H, Daniel S, Kusel K, Matthies C, Kuhner C, Braus-Stromeyer S (1997). ?Acetogenic bacteria: what are the in situ consequences of their diverse metabolic versatilities?”.
Biofactors
6
(1), 13?24. o.
PMID 9233536
.
- ↑
Morel, FMM, Kraepiel AML, Amyot M (1998). ?The chemical cycle and bioaccumulation of mercury”.
Annual Review of Ecological Systems
29
, 543?566. o.
- ↑
Dalton H (2005). ?
The Leeuwenhoek Lecture 2000 the natural and unnatural history of methane-oxidizing bacteria
”.
Philos Trans R Soc Lond B Biol Sci
360
(1458), 1207?22. o.
PMID 16147517
.
[
halott link
]
- ↑
Zehr J, Jenkins B, Short S, Steward G (2003). ?Nitrogenase gene diversity and microbial community structure: a cross-system comparison”.
Environ Microbiol
5
(7), 539?54. o.
PMID 12823187
.
- ↑
Koch A (2002). ?Control of the bacterial cell cycle by cytoplasmic growth”.
Crit Rev Microbiol
28
(1), 61?77. o.
PMID 12003041
.
- ↑
szerk.: Tor? Imre:
Az elet alapjai
. Budapest: Gondolat Kiado (1989).
ISBN 9632821815
- ↑
Eagon R. ?
Pseudomonas natriegens, a marine bacterium with a generation time of less than 10 minutes
”.
J Bacteriol
83
, 736?7. o.
PMID 13888946
.
- ↑
a
b
c
Thomson R, Bertram H (2001). ?
Laboratory diagnosis of central nervous system infections
”.
Infect Dis Clin North Am
15
(4), 1047?71. o.
PMID 11780267
.
- ↑
Paerl H, Fulton R, Moisander P, Dyble J. ?Harmful freshwater algal blooms, with an emphasis on cyanobacteria”.
ScientificWorldJournal
1
, 76?113. o.
PMID 12805693
.
- ↑
Challis G, Hopwood D. ?
Synergy and contingency as driving forces for the evolution of multiple secondary metabolite production by Streptomyces species
”.
Proc Natl Acad Sci U S A
100 Suppl 2
, 14555?61. o. [2008. julius 25-i datummal az
eredetib?l
archivalva].
PMID 12970466
. (Hozzaferes: 2006. december 29.)
- ↑
Kooijman S, Auger P, Poggiale J, Kooi B (2003). ?Quantitative steps in symbiogenesis and the evolution of homeostasis”.
Biol Rev Camb Philos Soc
78
(3), 435?63. o.
PMID 14558592
.
- ↑
Prats C, Lopez D, Giro A, Ferrer J, Valls J (2006). ?Individual-based modelling of bacterial cultures to study the microscopic causes of the lag phase”.
J Theor Biol
241
(4), 939?53. o.
PMID 16524598
.
- ↑
Hecker M, Volker U. ?General stress response of Bacillus subtilis and other bacteria”.
Adv Microb Physiol
44
, 35?91. o.
PMID 11407115
.
- ↑
Fraser C, Gocayne J, White O, Adams M, Clayton R, Fleischmann R, Bult C, Kerlavage A, Sutton G, Kelley J, Fritchman R, Weidman J, Small K, Sandusky M, Fuhrmann J, Nguyen D, Utterback T, Saudek D, Phillips C, Merrick J, Tomb J, Dougherty B, Bott K, Hu P, Lucier T, Peterson S, Smith H, Hutchison C, Venter J (1995). ?The minimal gene complement of Mycoplasma genitalium”.
Science
270
(5235), 397?403. o.
PMID 7569993
.
- ↑
Pradella S, Hans A, Sproer C, Reichenbach H, Gerth K, Beyer S (2002). ?Characterisation, genome size and genetic manipulation of the myxobacterium Sorangium cellulosum So ce56”.
Arch Microbiol
178
(6), 484?92. o.
PMID 12420170
.
- ↑
Hinnebusch J, Tilly K (1993). ?
Linear plasmids and chromosomes in bacteria
”.
Mol Microbiol
10
(5), 917?22. o.
PMID 7934868
.
- ↑
Brussow H, Canchaya C, Hardt W (2004). ?
Phages and the evolution of bacterial pathogens: from genomic rearrangements to lysogenic conversion
”.
Microbiol Mol Biol Rev
68
(3), 560?602. o.
PMID 15353570
.
- ↑
Perna N, Mayhew G, Posfai G, Elliott S, Donnenberg M, Kaper J, Blattner F (1998). ?
Molecular evolution of a pathogenicity island from enterohemorrhagic Escherichia coli O157:H7
”.
Infect Immun
66
(8), 3810?7. o.
PMID 9673266
.
- ↑
Denamur E, Matic I (2006). ?Evolution of mutation rates in bacteria”.
Mol Microbiol
60
(4), 820?7. o.
PMID 16677295
.
- ↑
Wright B (2004). ?Stress-directed adaptive mutations and evolution”.
Mol Microbiol
52
(3), 643?50. o.
PMID 15101972
.
- ↑
Davison J (1999). ?Genetic exchange between bacteria in the environment”.
Plasmid
42
(2), 73?91. o.
PMID 10489325
.
- ↑
Hastings P, Rosenberg S, Slack A (2004). ?
Antibiotic-induced lateral transfer of antibiotic resistance
”.
Trends Microbiol
12
(9), 401?4. o.
PMID 15337159
.
- ↑
a
b
c
Bardy S, Ng S, Jarrell K (2003). ?
Prokaryotic motility structures
”.
Microbiology
149
(Pt 2), 295?304. o. [2007. szeptember 14-i datummal az
eredetib?l
archivalva].
PMID 12624192
. (Hozzaferes: 2007. julius 1.)
- ↑
Wu M, Roberts J, Kim S, Koch D, DeLisa M (2006). ?
Collective bacterial dynamics revealed using a three-dimensional population-scale defocused particle tracking technique
”.
Appl Environ Microbiol
72
(7), 4987?94. o. [2008. szeptember 29-i datummal az
eredetib?l
archivalva].
PMID 16820497
. (Hozzaferes: 2006. december 30.)
- ↑
Lux R, Shi W (2004). ?Chemotaxis-guided movements in bacteria”.
Crit Rev Oral Biol Med
15
(4), 207?20. o.
PMID 15284186
.
- ↑
Frankel R, Bazylinski D, Johnson M, Taylor B (1997). ?
Magneto-aerotaxis in marine coccoid bacteria
”.
Biophys J
73
(2), 994?1000. o.
PMID 9251816
.
- ↑
Goldberg MB (2001). ?
Actin-based motility of intracellular microbial pathogens
”.
Microbiol Mol Biol Rev
65
(4), 595?626. o.
PMID 11729265
.
- ↑
E. Haeckel.
Generelle Morphologie der Organismen
. Reimer, Berlin (1866)
- ↑
E. Chatton.
Titres et travaux scientifiques
. Sette, Sottano, Italy (1937)
- ↑
H. F. Copeland.
The Classification of Lower Organisms
. Palo Alto: Pacific Books (1956)
- ↑
R. H. Whittaker (1969). ?New concepts of kingdoms of organisms”.
Science
163
, 150?160. o.
- ↑
C. R. Woese, W. E. Balch, L. J. Magrum, G. E. Fox and R. S. Wolfe (1977). ?An ancient divergence among the bacteria”.
Journal of Molecular Evolution
9
, 305?311. o.
- ↑
Woese C, Kandler O, Wheelis M (1990). ?
Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya.
”.
Proc Natl Acad Sci U S A
87
(12), 4576?9. o.
DOI
:
10.1073/pnas.87.12.4576
.
PMID 2112744
.
- ↑
Boucher Y, Douady CJ, Papke RT, Walsh DA, Boudreau ME, Nesbo CL, Case RJ, Doolittle WF (2003). ?Lateral gene transfer and the origins of prokaryotic groups.”.
Annu Rev Genet
37
, 283?328. o.
PMID 14616063
.
- ↑
Olsen G, Woese C, Overbeek R (1994). ?
The winds of (evolutionary) change: breathing new life into microbiology
”.
J Bacteriol
176
(1), 1?6. o.
PMID 8282683
.
- ↑
Woods G, Walker D (1996). ?
Detection of infection or infectious agents by use of cytologic and histologic stains
”.
Clin Microbiol Rev
9
(3), 382?404. o.
PMID 8809467
.
- ↑
Weinstein M (1994). ?
Clinical importance of blood cultures
”.
Clin Lab Med
14
(1), 9?16. o.
PMID 8181237
.
- ↑
Louie M, Louie L, Simor AE (2000). ?
The role of DNA amplification technology in the diagnosis of infectious diseases
”.
CMAJ
163
(3), 301?309. o.
PMID 10951731
.
- ↑
Oliver J. ?
The viable but nonculturable state in bacteria
”.
J Microbiol
43 Spec No
, 93?100. o. [2007. szeptember 28-i datummal az
eredetib?l
archivalva].
PMID 15765062
. (Hozzaferes: 2007. januar 22.)
- ↑
Barea J, Pozo M, Azcon R, Azcon-Aguilar C (2005). ?
Microbial co-operation in the rhizosphere
”.
J Exp Bot
56
(417), 1761?78. o.
PMID 15911555
. (Hozzaferes: 2006. december 31.)
- ↑
O'Hara A, Shanahan F (2006). ?The gut flora as a forgotten organ”.
EMBO Rep
7
(7), 688?93. o.
PMID 16819463
.
- ↑
Zoetendal E, Vaughan E, de Vos W (2006). ?A microbial world within us”.
Mol Microbiol
59
(6), 1639?50. o.
PMID 16553872
.
- ↑
Gorbach S (1990). ?
Lactic acid bacteria and human health
”.
Ann Med
22
(1), 37?41. o.
PMID 2109988
.
- ↑
Salminen S, Gueimonde M, Isolauri E (2005). ?
Probiotics that modify disease risk
”.
J Nutr
135
(5), 1294?8. o.
PMID 15867327
.
- ↑
Fish D. ?Optimal antimicrobial therapy for sepsis”.
Am J Health Syst Pharm
59 Suppl 1
, S13?9. o.
PMID 11885408
.
- ↑
Belland R, Ouellette S, Gieffers J, Byrne G (2004). ?Chlamydia pneumoniae and atherosclerosis”.
Cell Microbiol
6
(2), 117?27. o.
PMID 14706098
.
- ↑
Heise E. ?
Diseases associated with immunosuppression
”.
Environ Health Perspect
43
, 9?19. o.
PMID 7037390
.
- ↑
Saiman, L. ?Microbiology of early CF lung disease”.
Paediatr Respir Rev. volume = 5 Suppl A
, S367?369. o.
PMID 14980298
- ↑
Yonath A, Bashan A (2004). ?
Ribosomal crystallography: initiation, peptide bond formation, and amino acid polymerization are hampered by antibiotics
”.
Annu Rev Microbiol
58
, 233?51. o.
PMID 15487937
.
- ↑
Khachatourians G (1998). ?
Agricultural use of antibiotics and the evolution and transfer of antibiotic-resistant bacteria
”.
CMAJ
159
(9), 1129?36. o.
PMID 9835883
.
- ↑
Johnson M, Lucey J (2006). ?
Major technological advances and trends in cheese
”.
J Dairy Sci
89
(4), 1174?8. o.
PMID 16537950
.
- ↑
Hagedorn S, Kaphammer B. ?Microbial biocatalysis in the generation of flavor and fragrance chemicals”.
Annu Rev Microbiol
48
, 773?800. o.
PMID 7826026
.
- ↑
Cohen Y (2002). ?Bioremediation of oil by marine microbial mats”.
Int Microbiol
5
(4), 189?93. o.
PMID 12497184
.
- ↑
Neves L, Miyamura T, Moraes D, Penna T, Converti A. ?Biofiltration methods for the removal of phenolic residues”.
Appl Biochem Biotechnol
129?132
, 130?52. o.
PMID 16915636
.
- ↑
Liese A, Filho M (1999). ?Production of fine chemicals using biocatalysis”.
Curr Opin Biotechnol
10
(6), 595?603. o.
PMID 10600695
.
- ↑
Aronson A, Shai Y (2001). ?Why Bacillus thuringiensis insecticidal toxins are so effective: unique features of their mode of action”.
FEMS Microbiol Lett
195
(1), 1?8. o.
PMID 11166987
.
- ↑
Bozsik A (2006). ?Susceptibility of adult Coccinella septempunctata (Coleoptera: Coccinellidae) to insecticides with different modes of action”.
Pest Manag Sci
62
(7), 651?4. o.
PMID 16649191
.
- ↑
Chattopadhyay A, Bhatnagar N, Bhatnagar R (2004). ?Bacterial insecticidal toxins”.
Crit Rev Microbiol
30
(1), 33?54. o.
PMID 15116762
.
- ↑
Serres M, Gopal S, Nahum L, Liang P, Gaasterland T, Riley M (2001). ?
A functional update of the Escherichia coli K-12 genome
”.
Genome Biol
2
(9), RESEARCH0035. o.
PMID 11574054
.
- ↑
Walsh G (2005). ?Therapeutic insulins and their large-scale manufacture”.
Appl Microbiol Biotechnol
67
(2), 151 ? 9. o.
PMID 15580495
.
- ↑
Graumann K, Premstaller A (2006). ?Manufacturing of recombinant therapeutic proteins in microbial systems”.
Biotechnol J
1
(2), 164 ? 86. o.
PMID 16892246
.
- Ez a szocikk reszben vagy egeszben a
Bacteria
cim? angol Wikipedia-szocikk forditasan alapul. Az eredeti cikk szerkeszt?it annak laptortenete sorolja fel. Ez a jelzes csupan a megfogalmazas eredetet es a szerz?i jogokat jelzi, nem szolgal a cikkben szerepl? informaciok forrasmegjelolesekent.
Tovabbi informaciok
[
szerkesztes
]