Bakterie
(Bacteria, d?ive te? Bacteriophyta ?i Schizomycetes), nebo take
eubakterie
(Eubacteria), je
domena
jednobun??nych
prokaryotickych
organism?
. Mivaji
kokovity
?i
ty?inkovity
tvar a zpravidla dosahuji velikosti v ?adu n?kolika
mikrometr?
. Studiem bakterii se zabyva
bakteriologie
, vyznamn? tuto v?du rozvinuli
Robert Koch
a
Louis Pasteur
.
Typickou sou?asti bakterialnich bun?k je
peptidoglykanova
bun??na st?na
, jaderna oblast (
nukleoid
),
DNA
bez
intron?
,
plazmidy
a prokaryoticky typ
ribozom?
. U bakterii se nevyskytuje
pohlavni rozmno?ovani
, namisto toho se nej?ast?ji d?li
binarn?
. Bakterie jsou nejroz?i?en?j?i skupinou organism? na sv?t?. D?ive se druhy bakterii klasifikovaly podle vn?j?iho vzhledu, dnes jsou moderni zejmena
geneticke
metody. Diky nim se dnes rozli?uje asi 25 zakladnich
kmen?
bakterii.
Bakterie maji velky vyznam v
planetarnim ob?hu ?ivin
a mnohdy vstupuji do oboustrann? prosp??nych svazk? s jinymi organismy. Mnohe pat?i mezi
komenzalicke
druhy, ktere ?iji nap?iklad v lidske
travici soustav?
a n?ktere mohou byt dokonce symbioticke. Na druhou stranu je znamo i mnoho
patogennich bakterii
, tedy druh?, ktere zp?sobuji
infekce
. I ?lov?k mnohe z bakterii vyu?iva, nap?iklad v potravina?skem, leka?skem a chemickem pr?myslu. V?dci vyu?ivaji bakterie ve vyzkumu a samotne bakterie jsou p?edm?tem badani
bakteriologie
.
 Antoni van Leeuwenhoek
Antoni van Leeuwenhoek
byl prvni ?lov?k, ktery pozoroval bakterie
Bakterie poprve pozoroval roku
1676
nizozemsky p?irodov?dec
Antoni van Leeuwenhoek
, a to
mikroskopem
vlastni vyroby.
[1]
Jmeno
bacterium
zavedl a?
Christian Gottfried Ehrenberg
v roce
1828
[pozn. 1]
. Pojem pochazi z ?eckeho slova
bacterion
, ktere znamena maly klacek ?i ty?ka (prvni pozorovane bakterie byly
ty?inky
).
[3]
V roce 1859
Louis Pasteur
dokazal, ?e
kva?eni
zp?sobuji bakterie, a ?e tyto bakterie nevznikaji spontann? z ne?ive hmoty. Pasteur take prosazoval nazor, ?e mikroorganismy v?etn? bakterii zp?sobuji nemoci.
[4]
Robert Koch
byl pr?kopnikem v oblasti leka?ske mikrobiologie a studoval p?vodce
cholery
,
TBC
a
anthrax
. P?i vyzkumu TBC s kone?nou platnosti dokazal, ?e bakterie jsou p?vodci teto nemoci, za co? dostal v roce 1905
Nobelovu cenu
.
[5]
Takzvane
Kochovy postulaty
jsou vy?tem ?ty? kriterii, ktera jsou nutna k tomu, aby byl dany patogen uznan za p?vodce ur?ite nemoci.
V devatenactem stoleti ji? bylo znamo, ?e jsou mnohe bakterie patogenni, ale nebyly znamy u?inne antibakterialni leky.
[6]
V roce 1910 v?ak
Paul Ehrlich
vyvinul prvni
chemoterapeutikum
proti bakterii
Treponema pallidum
(p?vodce
syfilis
), a to diky zam?n? b??n? pou?ivanych laboratornich barviv za slou?eninu, ktera selektivn? zabijela bakterie.
[7]
Tradi?n? se v?ak za prvni systemov? pou?itelne
antibiotikum
uvadi
penicilin
, jeho? u?ink? si v?iml v roce
1928
Alexander Fleming
.
[8]
[9]
Gramovo barveni
, metoda k rychle klasifikaci bakterii do n?kolika skupin, bylo vyvinuto v roce 1884
Hansem Christianem Gramem
.
Spolu s vyvojem
bakteriologie
se vyvijelo i studium systematiky bakterii. Ve star?im pojeti zahrnovala ?i?e bakterie (
Monera
) v?echny
prokaryotni
organismy. V roce 1977
Carl Woese
zjistil, ?e
Archaea
maji vlastni vyvojovou linii, odli?nou od linie bakterii.
[10]
Z toho vyplyvajici taxonomie byla zalo?ena na sekvenaci ur?iteho useku
rRNA
, a rozd?lila prokaryoticke organismy na dv? samostatne domeny.
[11]
Bakterie jsou nejroz?i?en?j?i skupinou organism? na sv?t?.
[12]
Celkov? se odhaduje, ?e na Zemi ?ije asi 5×10
30
(jedinc?) bakterii. Celkovy po?et
druh?
se da jen tu?it, odhady sahaji od 10
7
k 10
9
druh?.
[13]
[14]
Bakterie je mo?ne nalezt v
p?d?
(jsou sou?asti
edafonu
a nejvice osidluji
rhizosferu
), ve
vod?
, v
ovzdu?i
i jako?to
symbionty
uvnit? a na povrchu
mnohobun??nych organism?
. V jednom gramu p?dy ?ije asi 40 milion? bakterii, v jednom
mililitru
sladke vody je jich p?ibli?n? milion.
[15]
Jsou v?ak znamy i druhy, ktere se specializuji na prost?edi, kde by ostatni organismy mohly p?e?ivat jen st??i (
vrouci
voda v
sope?nych jezerech
, nejvy??i vrstvy
atmosfery
a podobn?). N?ktere druhy bakterii jsou dle vyzkum? schopny p?e?it i ve
vesmiru
, tedy ve
vakuu
a o teplot? ?270
°C
.
[16]
Bakterie v?ak maji r?zne po?adavky na prost?edi v n?m? ?iji, vyznamnym hlediskem je
teplota
,
kyselost
a mno?stvi
kysliku
. Podle teplotniho optima se bakterie d?li na
psychrofilni
(do 20 °C),
mezofilni
(20?40 °C),
termofilni
(40 °C a vice) a p?ipadn? te?
hypertermofilni
s optimem kolem 80 °C.
[9]
Na zaklad? kyselosti rozli?ujeme
alkalofilni
(v zasaditem prost?edi),
neutrofilni
(v ± neutralnim prost?edi s pH 6?8
[9]
) ?i
acidofilni
(v kyselem prost?edi). Dal?im vyznamnym hlediskem je vztah ke kysliku.
Aerobni
bakterie (p?.
Mycobacterium
) vy?aduji kyslik v atmosfericke koncentraci,
mikroaerofilni
(
Lactobacillus
) v koncentraci velmi nizke (cca 2 %), v?t?ina bakterii v?ak pat?i mezi fakultativn?
anaerobni
, ktere rostou lepe v p?itomnosti kysliku, ale doka?ou r?st i bez n?ho. Na okraji tohoto spektra jsou striktn? anaerobni druhy, ktere ?iji jen v prost?edi bez kysliku (v?t?ina druh? rodu
Clostridium
).
[9]
Z dal?ich limitujicich ?initel? je mo?no zminit
vlhkost
(v?t?ina je vlhkomilnych, suchomilne jsou
nokardie
?i
aktinomycety
),
hydrostaticky tlak
(z hlubokych mo?i zname i
barofilni
bakterie),
osmoticky tlak
(p?edev?im v zavislosti na mno?stvi
soli
) a podobn?.
[9]
 Zakladni tvary bakterii
Zakladni tvary bakterii
Navenek je nejnapadn?j?im rysem bakterii tvar bakterialnich bun?k a jejich
kolonii
. U bakterii se rozli?uji nasledujici typy bun?k dle tvaru:
[17]
- kulovity
(
koky
) ? pokud vytva?eji kolonie, d?li se dale na diplokoky (kolonie tvo?ene dv?ma bu?kami), tetrakoky (?ty?i bu?ky v kolonii), streptokoky (?etizkovite kolonie), stafylokoky (hroznovite kolonie) a sarciny (bali?kovite kolonie)
- ty?inkovity
(
ty?inky
?ili bacily) ? mohou se sdru?ovat v koloniich po dvou (diplobacily) ?i v ?etizcich (streptobacily), p?ipadn? tvo?i palisady;
- zak?iveny
? takto tvarovane bakterie nevytva?eji kolonie a pat?i mezi n? vibria (kratke lehce zak?ivene ty?inky), spirily (lehce zvln?ne ty?inky) ?i spirochety (ty?inky ?rouboviteho tvaru);
- vlaknity
? vlaknite kolonie;
- v?tveny
? vytva?eji bu?to naznaky v?tveni nebo v?tveni uplne. Druha skupina m??e vytva?et bakterialni mycelia.
N?ktere bakterie vytva?eji kolonie podobne t?lu jednoduchych
mnohobun??nych
eukaryot
. Pat?i k nim n?ktere
sinice
(nap?.
Anabaena
),
myxobakterie
(Myxococcales) a mnohe dal?i skupiny. Druhe jmenovane bakterie dokonce tvo?i za ur?itych podminek makroskopicke
plodni?ky
se
sporami
uvnit?.
[18]
Take o dob?e znamych bakteriich, ktere napadaji lidske t?lo, se ukazuje, ?e doka?ou diky molekularnim signal?m synchronizovat sve chovani a ?tahnout za jeden provaz“.
[19]
[20]
[21]
Velikost bakterii je velice rozmanita, obvykle se ale pohybuje mezi desetinami a desitkami
mikrometr?
. Nap?iklad
Escherichia coli
dosahuje delky 2?3 μm a ?i?ky 0,6 μm.
[22]
Je v?ak znamo mnoho v tomto sm?ru extremnich bakterii.
D?ive se m?lo za to, ?e prokaryoticka bu?ka nem??e byt v?t?i ne? ?adov? desitky mikrometr? vzhledem k omezenym mo?nostem
difuze
, ktera hraje hlavni ulohu p?i transportu latek bu?kou. Mezi bakterie, ktere jsou z?ejm? vyjimkou z tohoto pravidla, pat?i nap?iklad
Epulopiscium fishelsoni
, jeji? 0,2?0,7 mm dlouhe bu?ky jsou v?t?i ne? bu?ky v?t?iny
prvok?
.
[23]
Do roku 2021 byla za nejv?t?i znamou bakterii pova?ovana
Thiomargarita namibiensis
(0,75 mm).
[24]
V r. 2022 v?ak byla popsana jeji p?ibuzna
Thiomargarita magnifica
?ijici v
karibskych
mangrovech
, ktera je o ?ad v?t?i ? dor?sta velikosti a? 2
cm
a v optimalnich podminkach by mohla byt i v?t?i. Navic je zajimava tim, ?e jeji genom je v bakterialni bu?ce obklopen membranovou strukturou.
[25]
[26]
Naopak nejmen?i jsou bakterie rodu
Mycoplasma
(a p?ibuzny
Ureaplasma
) s pr?m?rem bu?ky jen asi 0,1?0,3 μm. Nemaji
bun??nou st?nu
, a proto byly d?ive pova?ovany za
viry
.
[27]
Male jsou i
rickettsie
a
chlamydie
. Je?t? mnohem men?i jsou v?ak kontroverzni nalezy, ozna?ovane jako
nanobakterie
, ktere dosahuji velikosti jen 50?200
nanometr?
(0,05?0,2 μm),
[28]
[29]
podle nejnov?j?ich vyzkum? se v?ak nejedna o ?ive organismy, ale o krystaly
uhli?itanu vapenateho
v
krevnim seru
.
[30]
 Schema bakterialni bu?ky:
Schema bakterialni bu?ky:
A ?
pilus
, B ?
ribozom
, C ?
kapsula
, D ?
bun??na st?na
, E ?
bi?ik
, F ?
cytoplazma
, G ?
vakuola
, H ?
plazmid
, I ?
nukleoid
, J ?
cytoplazmaticka membrana
Bakterialni bu?ka je bu?kou
prokaryotni
. P?esto?e existuji zna?ne rozdily mezi stavbou bun?k r?znych skupin bakterii, lze vystopovat ur?ite spole?ne rysy. Zname nejen mnohe struktury na povrchu bun?k (
membrana
,
bun??na st?na
,
pilusy
,
bi?iky
), ale i vnitrobun??ne struktury (souhrnn?
protoplast
).
Na povrchu bakterialnich bun?k je
cytoplazmaticka membrana
podobna membran?
eukaryot
, ale s tim rozdilem, ?e v?t?inou neobsahuje ?adne
steroidy
. D?le?itou funkci membrany bakterii je tvorba
ATP
diky vytva?eni
protonoveho gradientu
(H
+
iont?
).
[27]
V?t?ina bakterii ma na povrchu bu?ky
bun??nou st?nu
z
peptidoglykanu
(murein), jen? obsahuje
kyselinu muramovou
jako jednu z hlavnich slo?ek. Peptidoglykan tvo?i kolem bun?k pevnou si? vypln?nou
peptidy
.
[17]
Bun??na st?na bakterii je v?ak velmi prom?nliva a je mimo jine d?le?itym znakem p?i rozli?ovani bakterii na
grampozitivni
a
gramnegativni
.
[27]
Grampozitivni maji toti? v bun??ne st?n? vice peptidoglykanu, ktery zachycuje vice
krystalove violeti
.
[31]
Vyjimku tvo?i bakterie t?idy
Mollicutes
(nap?. rod
Mycoplasma
), ktere nesyntetizuji peptidoglykan a tudi? jim chybi bun??na st?na. Jejich tvar tak dr?i pouze t?ivrstevna membrana, do ni? ukladaji steroidy. Diky teto zvla?tnosti maji tyto bakterie velkou plasticitu a mohou m?nit sv?j tvar.
[27]
[32]
 Pilusy
bakterie
Pilusy
bakterie
E. coli
slou?i
k p?ipevn?ni k podkladu
Mnoho bakterii ma na povrchu je?t? dal?i
polysacharidovou
?i
proteinovou
ochrannou vrstvu, tzv.
kapsulu
(pouzdro). Jeji funkci je dale zpev?ovat povrch bakterii (bakterie s kapsulou jsou diky tomu vice
patogenni
) a navic se lepe p?ichytit k substratu. N?kdy se diky
slizovitym
, men? soudr?nym kapsulam (zvanym
slizova vrstva
) va?ou jednotlive bu?ky v bakterialnich koloniich.
[27]
[31]
Dohromady se kapsule a slizove vrstv? take ?ika
glykokalyx
.
[27]
Vyjma plo?nych utvar?, ktere kryji bu?ku, se vyskytuje na povrchu je?t? ?ada jinych struktur, jako jsou nap?iklad
bi?iky
a
pilusy
(rovn?? zvane fimbrie
[33]
). Bi?iky jsou vlasovite (asi 20 nm silne, 20 μm dlouhe
[9]
) struktury ukotvene v membran?, tvo?ene
helikaln?
slo?enym proteinem
flagelinem
a slou?ici k pohybu.
[27]
Bakterie mohou mit ?adny, jeden (monotricha), vice na jedne stran? (lofotricha), dva uspo?adane proti sob? (amfitricha) ?i velky po?et bi?ik? rozlo?enych po celem povrchu bakterie (peritricha).
[34]
Bi?iky bakterii se stavbou zna?n? li?i od bi?ik?
eukaryot
.
Pilus
?ili fimbrie je druhym charakteristickym vlasovitym utvarem, ale krat?i, tu??i a u??i v pr?m?ru.
[33]
Pilusy bakterie vyu?ivaji k p?ichyceni na podklad (adheze), ale specializovane
sexualni pilusy
(F pilusy) slou?i k p?enosu
DNA
(sexualita bakterii).
[31]
Uvnit? bakterialni bu?ky se nachazi
protoplast
, tedy mno?stvi r?znych struktur rozptylenych v
cytosolu
. Mezi nejvyznamn?j?i vnitrobun??ne struktury u bakterii pat?i
nukleoid
(jaderne oblasti),
ribozomy
,
inkluze
a
cytoskelet
. V cytosolu v?ak jsou rozptyleny i jine struktury, nap?iklad mno?stvi
protein?
,
mRNA
a bakterialni
metabolity
.
Nukleoid (bakterialni chromozom, genofor) je jaderna oblast bakterii, tedy jediny obvykle kruhovy ?et?zec tvo?eny dv?ma vlakny
deoxyribonukleove kyseliny
(DNA). Vzacn? je v?ak DNA bakterii i linearni
[35]
nebo rozd?lena do vice chromozom?.
[36]
Nejsou p?itomny
histony
a netvo?i se ani
nukleozomy
, cela oblast ani zpravidla neni obklopena ?adnou membranou.
[27]
Op?t v?ak existuje vyjimka a nap?. u
planktomycet
a
poribakterii
byly nalezeny membranove struktury, ktere p?ipominaji eukaryoticke
bun??ne jadro
(u rodu
Gemmata
ma dokonce toto jadro dvojitou membranu a pory).
[37]
[38]
[39]
V cytoplazm? jsou pak roztrou?eny
plazmidy
, male useky DNA.
Ribozomy
jsou prokaryotickeho typu; jsou charakterizovany zkratkou 30
S
+50S, ktera vyjad?uje velikost a strukturu ribozomalnich podjednotek. Ribozomy bakterii jsou odli?ne od eukaryotickych nejen delkou protein?, ale i jejich typem, ?im? se logicky stavaji ?astym cilem antibiotik.
[27]
Inkluze jsou va?ky ?i zrna, zpravidla slou?ici jako zasobni struktury. Mohou skladovat
glykogen
,
fosfaty
, elementarni
siru
, n?kdy v?ak i
barviva
?i
enzymy
.
[33]
Bakterie pat?i k nejstar?im organism?m nalezanych ve form?
fosilii
. Prvni bakterie jsou znamy z
archaika
, nalezany jsou
stromatolity
, tedy horniny obsahujici fosilie bakterii, zvla?t?
sinic
.
[40]
Posledni spole?ny p?edek bakterii ale m??e byt star?i ne? 4 miliardy let.
[41]
P?edkem dne?nich bakterii byly jednobun??ne organismy, je? se vyvinuly na Zemi p?ed ?ty?mi miliardami let a pat?ily k v?bec prvnim formam
?ivota
. Dal?i t?i miliardy let v?echny organismy na Zemi byly mikroskopicke, p?i?em? po celou dobu dominovaly bakterie spolu s
archea
.
[42]
[43]
P?esto?e byly objeveny fosilie bakterii (nap?.
stromatolity
), nedostatek zjistitelnych znak? znemo??uje jejich ur?eni a hlub?i studium. Srozumitelne udaje se zji??uji pomoci
sekvenace
genomu recentnich bakterii. Diky teto technice v?dci ?aste?n? rekonstruovali bakterialni
strom ?ivota
a zda se, ?e bakterie jsou postranni v?tvi linie, z ni? se vyvinuly archea a
eukaryota
.
[44]
Poslednim spole?nym p?edkem
bakterii a archei byl pravd?podobn?
hypertermofil
, ktery ?il p?ed 2,5?3,2 miliardami let.
[45]
[46]
Bakterie se rovn?? zu?astnily vzniku
eukaryot
(viz
ni?e
).
 Znazorn?ni obrovske bohatosti bakterialni domeny (?edomod?e), ve srovnani eukaryotickymi organismy
Znazorn?ni obrovske bohatosti bakterialni domeny (?edomod?e), ve srovnani eukaryotickymi organismy
(?erven?) a domenou Archaea (zelen?)
Dnes se taxon bakterie (Bacteria) pova?uje za samostatnou
domenu
, ktera je striktn? odd?lena od ostatnich (domena
Archaea
i v?echny
eukaryoticke
?i?e). D?vodem k tomuto rozd?leni je skute?nost, ?e Archaea a Bacteria jsou velmi nep?ibuzne skupiny li?ici se velkym mno?stvim genetickych i morfologickych znak?.
Existuje vice zp?sob? klasifikace bakterii. Zpo?atku se klasifikovaly p?edev?im podle vzhledu (
fenotypu
), dnes se v?ak mimoto pou?ivaji te? analyticke (podle chemickych vlastnosti) a velmi ?asto te? geneticke (podle
genotypu
) metody.
[27]
Stejn? jako u jinych organizm? se pou?iva
binomicke nazvoslovi
(jako je nap?iklad
Escherichia coli
) a zakladnim
taxonem
je
druh
. Ni??imi taxony pak jsou
poddruh
, velmi ?asto i
morfovar
,
patovar
a
serovar
.
Ur?ovani (determinace, identifikace) bakterii ma velky vyznam v medicin?, kde je spravnym stanovenim p?vodce dane
bakterialni infekce
podmin?na nasledujici le?ba. Proto byla pot?eba identifikovat tyto bakterie hlavnim impulsem k vyvinuti determina?nich technik. Mikroskopickym pozorovanim t?lnich tekutin se bakterie ur?uji jen z?idka, ?ast?ji jsou preparaty barveny.
[9]
Znamym typem barveni je
Gramovo barveni
, ktere umo??uje rozli?ovat bakterie
grampozitivni
(G+),
gramnegativni
(G?) a
bez bun??ne st?ny
(Mollicutes). Pro
mykobakterie
(Mycobacteria) a
nokardie
(Nocardia) se zase pou?iva
Ziehlovo?Neelsenovo barveni
.
[49]
?asto v?ak nesta?i ani barvit vzorek, ale p?istupuje se ke kultivaci (viz
ni?e
). P?i identifikaci bakterii take ?im dal vice pou?iva take genetickych metod, jako je
polymerazova ?et?zova reakce
. Jejich vyhodou je jejich p?esnost a rychlost v porovnani s kultiva?nimi metodami.
[50]
Systematika se zabyva pojmenovavanim bakterialnich
taxon?
a jejich seskupovanim podle p?ibuznosti. Klasifikace bakterii je pr?b??n? vydavana v
International Journal of Systematic and Evolutionary Microbiology
(Mezinarodni ro?enka systematicke a evolu?ni mikrobiologie) a
Bergey's Manual of Systematic Bacteriology
(Bergeyho manual systematicke bakteriologie).
D?ive byla systematika zalo?ena p?edev?im na zaklad? morfologickych a analytickych metod, ktere v?ak dnes vytla?uji geneticke metody (podobn? jako v p?ipad? jejich ur?ovani). K pou?ivanym metodam pat?ilo i
Gramovo barveni
. Dal?imi mo?nostmi bylo d?leni na zaklad? rozdil? v bun??nem
metabolismu
, stavb? zakladnich bun??nych komponent (
DNA
,
mastnych kyselin
,
antigen?
apod.).
[51]
Tyto metody v?ak nezaru?uji p?irozenost taxon?, ktere byly na zaklad? nich vytvo?eny.
Dne?ni bakterialni klasifika?ni metody se soust?edi p?edev?im na molekularni systematiku. Z genetickych metod se vyu?iva
sekvenace
dlouhodob?ji stabilnich ?asti DNA, jako je
rRNA
nebo je m??en
obsah GC
(mno?stvi
guaninu
a
cytosinu
v ramci DNA).
[52]
Na zaklad? zmin?nych genetickych metod je v ramci domeny bakterie identifikovano pom?rn? velke mno?stvi
kmen?
. Jejich celkovy po?et stale p?ibyva; k b?eznu 2024 bylo platn? popsano 42 bakterialnich kmen? s kultivovanymi vzorky a navic 145 kmen? typu
Candidatus
, tedy bez uplneho popisu kv?li neexistenci kultivovanych vzork?.
[53]
[54]
[55]
P?ibuzne kmeny se seskupuji do vyvojovych linii (
klad?
), jednim ze znam?j?ich system? je ten
Cavalier-Smith?v
.
[56]
[57]
Od konce roku 2023 je pro vy??i seskupeni kmen? bakterii (i archei) po vzoru eukaryot oficialn? zavedena taxonomicka kategorie
?i?e
(
angl
.
kingdom,
lat
.
regnum).
[58]
Navr?eny jsou zatim ?ty?i bakterialni ?i?e:
[59]
[pozn. 2]
- Bacillati
Oren and Goker 2024
(d?ive uvad?na jako odd?leni
Firmicutes
a
Tenericutes
, resp. skupina
Terrabacteria
, resp. ?i?e
Terrabacterida
), zahrnujici kmeny
Actinomycetota
,
Armatimonadota
,
Bacillota
,
Cyanobacteriota
,
Chloroflexota
a
Mycoplasmatota
;
- Fusobacteriati
Battistuzzi and Hedges 2024
(d?ive uvad?na jako
Fusobacterida
), zahrnujici kmen
Fusobacteriota
;
- Pseudomonadati
(Gibbons and Murray) Oren and Goker 2024
(d?ive uvad?na jako odd?leni
Gracilicutes
, ?i?e
Hydrobacteria
resp.
Hydrobacterida
v?.
Aquificida
, resp. pod?i?e
Negibacteria
), zahrnujici kmeny
Abditibacteriota
,
Acidobacteriota
,
Aquificota
,
Atribacterota
,
Bacteroidota
,
Balneolota
,
Bdellovibrionota
,
Caldisericota
,
Calditrichota
,
Campylobacterota
,
Chlamydiota
,
Chlorobiota
,
Chrysiogenota
,
Coprothermobacterota
,
Deferribacterota
,
Desulfobacterota
,
Dictyoglomerota
,
Elusimicrobiota
,
Fibrobacterota
,
Gemmatimonadota
,
Ignavibacteriota
,
Kiritimatiellota
,
Lentisphaerota
,
Myxococcota
,
Nitrospinota
,
Nitrospirota
,
Planctomycetota
,
Pseudomonadota
,
Rhodothermota
,
Spirochaetota
,
Thermodesulfobacteriota
,
Thermodesulfobiota
,
Thermomicrobiota
,
Thermosulfidibacterota
a
Verrucomicrobiota
;
- Thermotogati
Battistuzzi and Hedges 2024
(d?ive uvad?na jako
Thermotogida
), zahrnujici kmeny
Deinococcota
,
Synergistota
a
Thermotogota
.
Bakterie v prost?edi, ktere jim zaji??uje pro n? vhodne chemicke i fyzikalni podminky, zpravidla
rostou
a
mno?i se
. Mechanismem r?stu je u bakterii
synteza
v?ech komponent t?la, ?im? se zvy?uje
hmotnost
i
objem
jedince. Po dosa?eni ur?itych rozm?r? se bakterie rozd?li metodou
binarniho d?leni
, co? je typ
nepohlavniho rozmno?ovani
. Doba mezi dv?ma d?lenimi se ozna?uje jako
genera?ni doba
. P?i dostate?nem po?tu bakterii v dane populaci lze mluvit o
kolonii
, doba pot?ebna k zdvojnasobeni po?tu bun?k v kolonii se nazyva
doba zdvojeni
.
[60]
Bakterie se rozmno?uji nej?ast?ji binarnim d?lenim, je znamo v?ak n?kolik p?ipad? odli?neho typu nepohlavniho mno?eni (nap?.
pu?eni
, tvorba
hormogonii
,
baeocyty
a podobn?).
P?i binarnim d?leni se bu?ka nejprve prodlou?i na dvojnasobnou delku a replikuje svou DNA, na?e? se uprost?ed za?ne vytva?et
septum
(p?ehradka slo?ena ze dvou
membran
a zakladu bun??ne st?ny). Septum v?dy vznika r?stem dvou p?epa?ek od protilehlych stran bu?ky do jejiho centra, kde se ob? ?asti spoji. P?i tomto procesu se uplat?uji r?zne enzymy (nap?.
transpeptidazy
). B??n? z ka?de mate?ske bu?ky vznikne jedna sesterska bu?ka, p?i nedokon?enem d?leni septa v?ak m??e dojit ke vzniku shluk? bakterii (viz ?lanek
koky
).
[27]
U
streptokok?
jsou mista r?stu vzajemn? orientovana pod uhlem 180°, ?im? vznikaji ?et?zce. U
stafylokok?
je tento uhel 90°, co? ma za nasledek vznik shluk?.
Vyjma p?i?neho d?leni je znamo je?t? mnoho jinych alternativnich zp?sob? nepohlavniho rozmno?ovani. Zvla?tnim zp?sobem je nap?iklad vytva?eni vice ne? jedne
spory
, ?im? se ze sporulace stava de facto rozmno?ovani. Takto se m??e za ur?itych podminek d?lit
Anaerobacter
(vznika najednou a? sedm spor). Dal?i mo?nosti je mnohonasobne d?leni jedne bu?ky na mnoho dil?ich
baeocyt?
u sinic ?adu
Pleurocapsales
. Mate?ska bu?ka obsahuje mnoho kopii DNA a v ur?item okam?iku prob?hne v rychlem sledu mnohonasobne d?leni a z popraskane mate?ske bu?ky se uvolni nove bu?ky. Naopak pu?eni, ktere se vyskytuje nap?iklad u
sinic
nebo u
Planctomycetes
, probiha tak, ?e zrala bu?ka za?ne na svem povrchu vytva?et zcela novou bu?ku. Pokud dce?ina bu?ka doroste do dostate?ne velikosti, je do ni vpu?t?na DNA a bu?ka (obvykle podstatn? men?i ne? mate?ska) je odpojena. Postupem ?asu doroste a m??e se za?it sama rozmno?ovat. I zde p?edchazi pu?eni
replikace DNA
.
[61]
N?ktere p?isedle bakterie (nap?. sinice) se mno?i pomoci
hormogonii
, kratkych vlaken, ktera se odd?li od mate?skeho ?et?zce a dor?staji v novy.
[62]
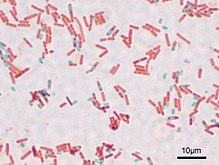 Vznik
endospor
u bakterii rodu
Bacillus
; na teto fotografii jsou spory obarveny zelen?, zatimco vegetativni bu?ky kolem nich jsou ?ervene
Vznik
endospor
u bakterii rodu
Bacillus
; na teto fotografii jsou spory obarveny zelen?, zatimco vegetativni bu?ky kolem nich jsou ?ervene
(
sv?telny mikroskop
)
N?ktere bakterie vytva?eji
spory
, tedy zvla?tni bu?ky, ktere slou?i k dlouhodobemu p?e?iti nep?iznivych podminek. Takove bakterie nazyvame
sporulujici
. Spory tvo?i zejmena n?ktere
grampozitivni bakterie
- bakterie ze skupiny
Firmicutes
(modelove rody
Clostridium
?i
Bacillus
). Jejich spory nazyvame
endospory
, proto?e vznikaji uvnit? bu?ky mate?ske, a to v?dy jen jedna v ka?de bu?ce, zatimco okolni bu?ka mate?ska se posleze rozpusti.
[60]
Jejich schopnost vydr?et nehostinne podminky se ozna?uje jako
kryptobioza
. V praxi mohou spory p?e?it extremn? vysoke
teploty
(n?kdy vydr?i a? n?kolik hodin
varu
),
radiaci
,
kyselost
prost?edi, dezinfek?ni latky v prost?edi a podobn?. Tim se bakterialni spory pravd?podobn? stavaji nejodoln?j?imi znamymi bu?kami v p?irod?. Endospory se za p?izniv?j?ich okolnosti op?t zm?ni ve vegetativni bu?ky.
[33]
- Spory odli?neho typu vytva?i jine, take grampozitivni bakterie, konkretn?
aktinomycety
. Ty vznikaji na konci vlakna aktinomycet, a proto se nejedna o endospory, a navic maji mirn? odli?ne vlastnosti.
- T?eti skupinou, je? sporuluje, jsou bakterie rodu
Azotobacter
, v tomto p?ipad? se v?ak odolne utvary spi?e nazyvaji
cysty
. Cysty maji zpomaleny
metabolismus
a ztloustlou bun??nou st?nu.
[60]
Souvisejici informace naleznete take v ?lanku
?ivna p?da
.
 Petriho misky
s
krevnim agarem
Petriho misky
s
krevnim agarem
V
laborato?i
se mnohdy z r?znych d?vod? bakterie kultivuji v um?lem prost?edi (
mediu
). Toto prost?edi musi mit vhodne chemicke a fyzikalni vlastnosti, zpravidla charakteristicke pro ur?ity typ bakterii. Medium musi proto obsahovat v?echny nezbytne
?iviny
nutne pro ?ivot a r?st bakterii. Ka?da bakterie po?aduje ur?ite
mineralni latky
, ty se nej?ast?ji p?idavaji ve form? r?znych slou?enin.
[63]
Kultiva?ni media pro?la od dob
Pasteura
zna?nym vyvojem, jako ?ivna p?da pro mikroorganismy se pou?ivaly vyvary z
kvasnic
,
komorova voda
z o?i, pozd?ji
agar
(
polysacharid
z
ruduch
). V klinicke
bakteriologii
je dnes zakladem v?t?iny ?ivnych medii
krevni agar
p?ipravovany smi?enim
ov?i
krve
a
agaru
. M??e byt dale upravovan pro ur?ite skupiny bakterii (nap?.
?okoladovy agar
pro
meningokoky
,
?ulova p?da
pro
mykobakterie
). Takzvana
selektivni media
se pou?ivaji v p?ipad?, kdy je pot?eba ziskat
?istou kulturu
ur?itych bakterii ? nap?iklad po p?idani 10% roztoku
chloridu sodneho
vyp?stujeme kulturu
stafylokok?
.
Diagnosticke medium
umo??uje diagnostikovat ur?ity druh bakterii, nap?. podle jejich
metabolickeho produktu
.
[64]
V?t?ina bakterii obsahuje jediny
nukleoid
(tzv. bakterialni
chromozom
), obvykle kruhovou molekulu
DNA
, ktera se sklada z
nukleovych bazi
. Vyjimkou jsou nap?.
spirochety
z rodu
Borrelia
majici nukleoid linearniho (nikoliv kruhoveho) tvaru.
[35]
[65]
Po?et bazi saha od 112 000
komplementarnich par?
bazi (u
Nasuia deltocephalinicola
)
[66]
a? k 12 200 000 par?m u p?dni bakterie
Sorangium cellulosum
.
[67]
Geny
jsou v?t?inou po cele delce DNA, p?esto?e i u n?kterych bakterii existuji
introny
(
nep?ekladane
?asti gen?).
[68]
Dale bakterie obsahuji
plazmidy
, izolovane ?asti DNA, obsahujici v?dy jen n?kolik malo
gen?
. Bakterie se bez nich sice zpravidla obejdou, plazmidy v?ak slou?i bakteriim nap?iklad pro rezistenci k
antibiotik?m
, k fixaci vzdu?neho
dusiku
a k jinym specializovanym u?el?m, ?im? svym hostitel?m poskytuji selek?ni vyhodu. Plazmidy se mohou mezi bakteriemi horizontaln? p?ena?et procesy
transformaci
,
konjugaci
a
transdukci
.
[31]
Geny jsou v procesu
transkripce
p?episovany do jednotlivych typ?
RNA
, jako je
mRNA
,
rRNA
?i
tRNA
. Z mRNA se pak syntetizuji proteiny v procesu
translace
. Bakterie se rozmno?uji nepohlavn?, a proto
d?di
identicke kopie genom? svych rodi?? (jsou to
klony
). P?esto se DNA m??e vyvijet diky
rekombinaci
?i
mutacim
. Mutace jsou zp?sobeny chybami p?i
replikaci DNA
a vystavenim r?znym
mutagen?m
. Mezi bakteriemi dochazi k vym?n? genetickeho materialu
horizontalnim p?enosem
.
Bakterie nemaji system pohlavi podobny
eukaryotickym organism?m
, nicmen? mohou si navzajem vym??ovat ?ast svoji geneticke informace. Tento proces se ozna?uje jako
konjugace
. P?enos probiha jednim sm?rem: jedna z bakterii je darce ?ili donor DNA (nespravn? take ?sam?i bu?ka“), druhy je p?ijemce ?ili recipient (?sami?i bu?ka“). Vym?na DNA se d?je spojenim bun?k p?es
sexualni pilusy
, vlaknite dute utvary. Schopnost tvo?it sexualni pilusy je umo?n?na specifickymi sekvencemi DNA, je? se souhrnn? ozna?uji jako
F-faktor
. V praxi m??e byt F-faktor jak ?ast
bakterialniho chromozomu
, tak
plazmid
. Darce geneticke informace musi mit F-faktor, proto je zvan F
+
.
[31]
Konjugaci se zvy?uje geneticka diverzita bakterii.
Bakterie mohou geneticky material p?ijimat i z okolniho prost?edi, nap?. z mrtvych bakterii. Tento proces se ozna?uje jako
transformace
. Proces
transdukce
zase v praxi znamena p?enos genetickeho materialu bakterialnimi
viry
(
bakteriofagy
).
[31]
 Vlakna sinic obsahuji
Vlakna sinic obsahuji
fotosynteticka barviva
a probiha
u nich oxygenni
fotosynteza
(
sv?telny mikroskop
)
U bakterii je znamo velke mno?stvi r?znych
metabolickych
proces?, pravd?podobn? mnohem vic ne? u
eukaryotickych
organism?.
[69]
Tradi?n? byl typ metabolismu jednim ze zakladnich znak? pro taxonomii bakterii, ale dnes se vi, ?e taxonomie zalo?ena na t?chto znacich ?asto neodpovida skute?nemu
fylogenetickemu vyvoji
.
[70]
Metabolicka aktivita je samoz?ejm? podmin?na obsahem biogennich prvk? v substratu (uhlik, dusik, sira, fosfor) a vhodneho zdroje
energie
k
biosyntetickym
proces?m.
Auxotrofni
druhy, tedy bakterie, ktere si neumi ur?itou slou?eninu syntetizovat, vy?aduji te? n?ktere
r?stove faktory
(
vitaminy
,
aminokyseliny
,
purinove
a
pyrimidinove
baze).
[71]
Bakterialni metabolismus m??eme rozd?lit podle t?i kriterii: druh u?ivane
energie
, zdroj
uhliku
a
donor
elektron?
.
[72]
Dle druhu u?ivane energie se rozli?uji bakterie
fototrofni
a
chemotrofni
, podle zdroje uhliku na
autotrofni
a
heterotrofni
a dle donor? elektron? na
litotrofni
a
organotrofni
. Tyto terminy jsou vzajemn? kombinovatelne, proto nap?iklad
cyanobakterie
jsou
fotoautotrofni
.
K fototrofnim bakteriim pat?i p?edev?im ji? zmin?ne
cyanobakterie
(sinice), dale
zelene sirne bakterie
,
zelene nesirne bakterie
,
heliobacterie
,
purpurove bakterie
[73]
a v r. 2007 popsane
chloracidobakterie
.
[74]
V?t?ina se ozna?uje za
fotoautotrofy
, proto?e zpravidla ziskavaji uhlik z anorganickych zdroj? (
oxid uhli?ity
). U t?chto bakterii probiha
fotosynteza
, proces, ktery pohani energie ze slune?niho sv?tla a jeho vysledkem je fixace uhliku v organickych slou?eninach za pomoci
Calvinova cyklu
. Donorem elektron? pro redukci
NADP
+
je bu?
voda
(tzv. oxygenni fotosynteza, uvolni se
kyslik
), nebo jine latky (
vodik
,
sira
,
thiosulfat
,
sirovodik
), p?i nich? se kyslik neuvol?uje. Sinice vyu?ivaji p?i fotosynteze barviva
chlorofylu
a dal?ich pigment?. Mimo fotoautotrofy existuje i n?kolik
fotoheterotrof?
, ktere sice take fotosyntetizuji, ale jako zdroje uhliku u?ivaji organickych slou?enin (
acetat
,
pyruvat
).
[71]
Chemotrofni bakterie, tedy bakterie vyu?ivajici energie z vhodnych chemickych reakci, mohou byt rovn?? dvojiho typu. Takzvane
chemoautotrofni
ziskavaji uhlik z anorganickych slou?enin, typicky oxidu uhli?iteho a
chemoheterotrofni
z organickych slou?enin. K chemoheterotrofnim bakteriim ?adime nap?iklad
rozklada?e
v p?d? (v podstat?
saprofyty
) nebo bakterie podilejici se na
kvasnych
procesech. Zname v?ak rovn?? mnoho chemoautotrofnich skupin, nap?iklad bakterie
vodikove
,
sirne
, bakterie oxidujici
?elezo
?i siru,
nitrifika?ni
a dal?i.
[71]
Bakterie diky svym unikatnim metabolickym pochod?m hraji vyznamnou roli v kolob?hu latek, a to zejmena v kolob?hu
uhliku
,
dusiku
a
siry
. Tim, ?e se podileji na
rozkladu
organickych latek, vraci do atmosfery
oxid uhli?ity
. N?ktere bakterie doka?ou z atmosfery
fixovat vzdu?ny dusik
, jako nap?iklad
Rhizobium
, ?i sinice
Anabaena
a
Nostoc
. N?ktere z t?chto bakterii jsou rovn?? obsa?eny v hlizkach
bobovitych
rostlin. Vyznamne jsou i bakterie, zapojene v p?em?n? dusikatych latek na
dusitany
a
dusi?nany
, p?ipadn? zp?t na vzdu?ny dusik.
[75]
 Mitochondrie
vznikly pravd?podobn? z
endosymbiotickych
bakterii
Mitochondrie
vznikly pravd?podobn? z
endosymbiotickych
bakterii
(
transmisni elektronovy mikroskop
)
Bakterie jsou v p?irod? nezastupitelne ve svem vyznamu pro
kolob?h latek
, jako symbioticke
oboustrann? prosp??ne
organismy ?i jako vyrobni prost?edek v
biotechnologiich
. Dale jsou tu take bakterie zp?sobujici choroby a bakterie podilejici se na rozkladu mrtve organicke hmoty (
destruenti
). Hydrolyticke bakterie jsou zodpov?dne za
hydrolyzu
organickych latek.
Velmi vyznamn? se v historii bakterie zapojily do vzniku
eukaryotickych bun?k
. Bakterie maji i sve patogeny, a to p?edev?im
viry
, ktere se nazyvaji
bakteriofagy
.
Podle endosymbioticke teorie se davne bakterie podilely na vzniku
eukaryoticke bu?ky
z primitivnich bun?k, ktere mohly byt p?ibuzne dne?nim archeim.
[76]
[77]
Alfaproteobakterie
byly pohlceny proto
eukaryotickymi bu?kami
za vzniku
mitochondrii
?i
hydrogenozom?
. Tyto
organely
se stale nachazi u v?ech znamych eukaryot (n?kdy v redukovane podob?). Pozd?ji vedlo pohlceni dal?ich endosymbiont?, tentokrat bakterii p?ibuznych
sinicim
, ke vzniku
chloroplast?
?as
a
rostlin
.
[78]
[79]
Je znamo velke mno?stvi
mutualistickych
(oboustrann? prosp??nych) symbiotickych vztah? bakterii. Velmi ?asto se v symbiotickych svazcich u?astni
sinice
. Ty vstupuji jak svazk? s
houbami
(tzv.
cyanoli?ejniky
), ale velmi ?asto te? tvo?i symbioticke vztahy s
vy??imi rostlinami
(sinice jako
fixato?i dusiku
). Symbioticke sinice vegetujici v t?lech
?ivo?ich?
se nazyvaji
zoocyanely
a maji je nap?iklad
pla?t?nci
(Tunicata).
[80]
Mimo sinic je znamo i mnoho heterotrofnich bakterii, ktere se u?astni symbiotickych svazk?. Vyznamne jsou p?edev?im
hlizkove bakterie
(nap?.
Rhizobium
) a take mno?stvi bakterii v t?lnich dutinach ?ivo?ich? (nap?iklad jako sou?ast
st?evni mikroflory
). Znamy jsou v?ak i symbioticke
bioluminiscen?ni
bakterie, bakterie
travici celulozu
a mnohe jine.
[80]
 Bacteroides
Bacteroides
je nej?ast?j?i lidska
st?evni bakterie
[81]
(
sv?telny mikroskop
)
Na povrchu i uvnit? t?l organism? ?ije pom?rn? zna?ne mno?stvi bakterii, ktere nazyvame
komenzalicke
, pokud svemu
hostiteli
vyrazn? ne?kodi, ale ani vyrazn? neprospivaji. V lidskem t?le se vyskytuje
mikrobiom
na
k??i
, v cele
travici soustav?
(zejmena v
tlustem st?ev?
), v hornich
dychacich cestach
, v
uchu
a
oku
,
mo?ove trubici
a
vagin?
(
vaginalni flora
).
[27]
Takzvana
st?evni mikroflora
mnoha obratlovc? v?etn? ?lov?ka je p?eva?n? tvo?ena prav? komenzalickymi bakteriemi, ktere jsou v
tlustem st?ev?
p?itomny v obrovskych po?tech. Mno?stvi bakterii v gramu st?evni
traveniny
se odhaduje na 10
12
a p?edpoklada se, ?e celkove mno?stvi st?evnich bakterii dosahuje po?tu lidskych
bun?k
daneho jedince ?i ho dokonce p?evy?uje.
[82]
[83]
N?kdy komenzalicke bakterie p?echazi v
patogeny
, nebo se naopak stavaji
mutualisty
.
[84]
Mno?stvi bakterii vytva?i
mikrobialni
povlak i na lidske
k??i
. P?esto?e je lidska
poko?ka
sucha, slana a
kysela
, roste na ni velke mno?stvi bakterii, jako
Corynebacterium
,
Staphylococcus
,
Micrococcus
a mnohe dal?i. V?t?inou ko?ni bakterie ?iji v blizkosti
potnich ?laz
a u ko?ene
vlas?
. Bakterie jsou i d?vodem, pro? lide pachnou, kdy? se
poti
.
[85]
N?ktere bakterie zp?sobuji
choroby
, n?kdy souhrnn? zvane
bakteriozy
?i bakterialni infekce. Ty se neomezuji na ?lov?ka, naopak r?zne bakterie napadaji ?iroke spektrum hostitelskych druh? v?etn?
hub
,
rostlin
a
prvok?
. P?esto?e n?ktere bakterie mohou byt patogeny i na samotnem povrchu t?la daneho organismu, zna?ne mno?stvi jich vstupuje dovnit? r?znymi t?lnimi otvory, nap?iklad u rostlin
pr?duchy
, skrz
sliznice
?ivo?ich?,
ranami
, p?ipadn? p?es
k??i
. V mist?, kde se usidli, mohou zp?sobit
hnisani
, ni?it tka? ?i pletiva (nap?.
nekroza
) ?i ?kodit svymi vlastnimi
toxiny
.
Nazvy bakterialnich nemoci se ?asto tvo?i p?idanim koncovky -oza k nazvu dane patogenni bakterie. Mezi nejb??n?j?i lidske bakterialni nemoci pat?i nap?iklad
zubni kaz
, z va?n?j?ich nemoci je ve sv?t? velmi ?asta
tuberkuloza
, v roce 2002 ji byly podle
WHO
infikovany dv? miliardy lidi a ro?n? na ni umiraly dva miliony lidi.
[27]
Existuji r?zne mo?nosti nakazy. S potravou (alimentarn?) se do t?la dostava nap?iklad
Salmonella
(
salmoneloza
),
Shigella
spp. (
shigeloza
provazena
pr?jmy
) nebo
Listeria
spp. (p?vodce
listeriozy
), vzdu?nou cestou nap?iklad
Mycobacterium
spp. (
TBC
a
lepra
) nebo
Chlamydophila psittaci
(p?vodce jedne z
chlamydioz
). Ranami se do t?la dostane nap?iklad p?vodce
tetanu
,
Clostridium tetani
. Specialni p?ipad jsou p?enosy p?es
?lenovce
, vyskytujici se nap?iklad u bakterii
Borrelia
spp. (jeden druh p?vodcem
boreliozy
),
Rickettsia
spp. (
tyfus
a jine), a u mnohych dal?ich. P?es urogenitalni trakt se bakterie mohou dostat do t?la
pohlavnim stykem
, jako nap?iklad
Neisseria gonorrhoeae
(
kapavka
) ?i
Treponema pallidum
(
syfilis
).
[27]
Lidske t?lo bojuje s bakteriemi pomoci n?kterych slo?ek imunitniho systemu. Proti extracelularnim bakteriim (?ijicim mimo lidske bu?ky, obvykle infekce) v t?le bojuji hlavn? bu?ky
neutrofily
? k tomu jim v?ak pomaha i tzv.
komplement
a r?zne
protilatky
. Vnitrobun??ne bakterie jsou cilem aktivovanych
makrofag?
a
T
C
-lymfocyt?
.
[86]
?lov?k sam ma v sob? i na sob? r?zne druhy bakterii, bez kterych by se ?asto neobe?el (
symbioza
).
[87]
Tyto jsou nazyvany souhrnn? jako lidsky
mikrobiom
. Diverzita
patogen?
je v?t?i u domorodc?,
[88]
kte?i nep?i?li do styku s moderni le?bou (ktera zp?sobuje stavy jako je
antibioticka rezistence
).
Bakterie, jako
Lactobacillus
, se (spole?n? s
kvasinkami
a
plisn?mi
) ?asto ji? tisice let pou?ivaji k p?iprav?
fermentovanych
(kva?enych) potravin, jako je
syr
,
sojova oma?ka
, nakladana
zelenina
,
kysele zeli
,
ocet
,
vino
a
jogurt
.
[89]
[90]
Jogurt a kefir se vyrabi kva?enim mleka za p?itomnosti bakterii; mleko diky tomuto procesu dostava jine p?ichuti.
[17]
Mle?nych bakterii se vyu?iva p?i pr?myslove produkci
kyseliny mle?ne
. Ta toti? vznika kva?enim
sacharid?
. Fermentaci sacharid? za p?itomnosti maselnych bakterii se zase vyu?iva k produkci
kyseliny octove
a
maselne
, kva?enim vznikle kyseliny maselne se pr?myslov? vyrabi
butanol
a
aceton
.
[17]
Dal?imi latkami, je? jsou produkovany bakteriemi, jsou
xanthan
(pou?iva se jako mazivo, p?isada do potravin, p?i vyrob? nat?rovych hmot, keramiky a r?znych dal?ich prost?edk?) a
kurdlany
(potencialni uplatn?ni v potravina?stvi).
[91]
V pr?myslu se te? uplat?uji
enzymy
ziskane z bakterii. N?ktere
proteazy
se p?idavaji do n?kterych
pracich pra?k?
, ke ?t?peni ?krobu se u?iva
amylaz
, v medicin? na?ly uplatn?ni
streptokinazy
.
[9]
Mnohdy se v biotechnologii p?istupuje k cilenemu ?lecht?ni bakterii za u?elem zlep?eni jejich vlastnosti. V praxi to znamena hledat v kultu?e nahodne
mutanty
, bakterie s, nebo bez ur?iteho
plazmidu
, nebo je cilen?
rekombinovat
a mutovat. Uva?uje se nap?iklad o ?lecht?ni bakterii
fixujicich dusik
.
[92]
Vyu?iti v ochran? ?ivotniho prost?edi
[
editovat
|
editovat zdroj
]
Schopnost bakterii rozkladat mnohe latky se vyu?iva p?edev?im v zpracovavani (nap?. toxickeho)
odpadu
i jinych zp?sobech
bioremediace
. V
?istirnach odpadnich vod
je velmi ?asto podporovan r?st
aerobnich
rozkladnych bakterii tim, ?e je odpadni voda promichavana kyslikem za vzniku tzv.
aktivovaneho kalu
,
[93]
rozkladnych bakterii se v?ak na podobnem principu vyu?iva i v r?znych domacich
septicich
.
[94]
Bakterie schopne travit
uhlovodiky
v
rop?
jsou vyu?ivany p?i ?i?t?ni
ropnych skvrn
,>
[95]
na pla?e se n?kdy p?idava hnojivo, aby se r?st bakterii podpo?il (nap?. po havarii tankeru
Exxon Valdez
). V chemickem pr?myslu se bakterie pou?ivaji k produkci nejr?zn?j?ich
chemickych latek
, p?ipadn?
lek?
?i
agrochemikalii
.
[96]
Bakterie se rovn?? pou?ivaji namisto
pesticid?
v
biologickem boji
proti
?k?dc?m
. V tomto ohledu je znama p?dni bakterie
Bacillus thuringiensis
(BT).
[97]
Pomoci
geneticky upravenych
bakterii se take vyrabi
inzulin
a dal?i
hormony
,
enzymy
,
r?stove faktory
?i
protilatky
.
[98]
[99]
Ve vyzkumu se bakterii vyu?iva kv?li rychlemu r?stu a pom?rn? snadne manipulaci s nimi. Bakterie jsou modelove organismy pro
molekularni biologii
,
genetiku
a
biochemii
. V?dci nap?iklad cilen?
mutuji
DNA bakterii a nasledne
fenotypy
zkoumaji ? timto zp?sobem se zji??uje funkce
gen?
,
enzym?
a
metabolickych cest
, jejich zna?na ?ast se da pozd?ji aplikovat i na komplexn?j?i organismy.
[100]
Modelovym organismem je zejmena bakterie
Escherichia coli
.
[101]
Schopnosti n?kterych patogennich bakterii inkorporovat plazmidy do DNA hostitele se vyu?iva v genetickem in?enyrstvi: zejmena
Agrobacterium tumefaciens
je pou?ivana p?i cilene p?iprav?
geneticky modifikovanych plodin
.
[102]
Pro velice d?le?itou vyzkumnou a diagnostickou metodu
PCR
je nutna termostabilni
DNA polymeraza
. Nej?ast?ji vyu?ivane polymerazy pro PCR jsou odvozeny z polymerazy termofilni bakterie
Thermus aquaticus
.
[103]
Bakterialniho p?vodu je te? system
CRISPR-Cas
?iroce vyu?ivany pro editaci DNA.
[104]
- ↑
jako
rodove
jmeno ve sve ustni p?edna?ce, tiskem publikovane a? v r. 1830; nominativ pluralu
Bacteria
byl poprve pou?it a? v r. 1845
[2]
- ↑
U navr?enych ?i?i jsou v zahrnutych kmenech uvad?ny jen kmeny s platn? publikovanymi nazvy, nikoli kmeny typu
Candidatus
a jine dosud neplatn? popsane kmeny, a? by do ?i?i z fylogenetickeho hlediska pravd?podobn? spadaly
- ↑
PORTER, J R. Antony van Leeuwenhoek: tercentenary of his discovery of bacteria.. S. 260?269.
Bacteriological Reviews
[online]. 1976-06 [cit. 2021-01-06]. Ro?. 40, ?is. 2, s. 260?269.
Dostupne online
.
PMID
786250
. (anglicky)
- ↑
SH?SHKIN-SKARÐ, Yegor. When and how did the names Bacteria and Eubacteria originate: Resurrected facts.
Microbiological Research
[online]. G. Fischer in Elsevier GmbH., 2024-06 [cit. 2024-04-11]. Ro?. 283: 127676.
Dostupne online
.
ISSN
1618-0623
.
DOI
10.1016/j.micres.2024.127676
.
PMID
38503219
. (anglicky)
- ↑
Online Etymology dictionary.
Etymology of the word "bacteria"
[online].
Dostupne online
. (anglicky)
- ↑
Pasteur's Papers on the Germ Theory
[online]. LSU Law Center's Medical and Public Health Law Site, Historic Public Health Articles.
Dostupne online
.
- ↑
The Nobel Prize in Physiology or Medicine 1905
[online]. Nobelprize.org.
Dostupne online
. (anglicky)
- ↑
THURSTON, Alan J. Of Blood, Inflammation and Gunshot Wounds: The History of the Control of Sepsis. S. 855?861.
ANZ Journal of Surgery
[online]. 2000-12-05 [cit. 2021-01-06]. Ro?. 70, ?is. 12, s. 855?861.
Dostupne online
.
DOI
10.1046/j.1440-1622.2000.01983.x
. (anglicky)
- ↑
SCHWARTZ, Robert S. Paul Ehrlich's Magic Bullets. S. 1079?1080.
New England Journal of Medicine
[online]. 2004-03-11 [cit. 2021-01-06]. Ro?. 350, ?is. 11, s. 1079?1080.
Dostupne online
.
DOI
10.1056/NEJMp048021
. (anglicky)
- ↑
KLABAN, Vladimir.
Sv?t mikrob?; ilustrovany lexikon mikrobiologie ?ivotniho prost?edi
. Hradec Kralove: Gaudeamus, 2001.
ISBN
80-7041-687-4
. S. 416.
- ↑
a
b
c
d
e
f
g
h
VOTAVA, Miroslav.
Leka?ska mikrobiologie obecna
. [s.l.]: Neptun, 2001.
- ↑
WOESE, C. R.; FOX, G. E. Phylogenetic structure of the prokaryotic domain: The primary kingdoms. S. 5088?5090.
Proceedings of the National Academy of Sciences
[online]. 1977-11-01 [cit. 2021-01-06]. Ro?. 74, ?is. 11, s. 5088?5090.
Dostupne online
.
DOI
10.1073/pnas.74.11.5088
.
PMID
270744
. (anglicky)
- ↑
WOESE, Carl R.; KANDLER, Otto; WHEELIS, Mark L. Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya.. S. 4576?4579.
Proceedings of the National Academy of Sciences
[online]. 1990-06-01 [cit. 2021-01-06]. Ro?. 87, ?is. 12, s. 4576?4579.
Dostupne v archivu
po?izenem z
originalu
dne 2008-06-27.
DOI
10.1073/pnas.87.12.4576
.
PMID
2112744
. (anglicky)
- ↑
LAWTON, Wendy Y.
Researchers Break New Ground in Their Study of Bacteria
[online]. Inside Brown [cit. 2008-04-22].
Dostupne v archivu
po?izenem dne 06-04-2008. (anglicky)
? neplatny odkaz !
- ↑
CURTIS, Thomas P.; SLOAN, William T.; SCANNELL, Jack W. Estimating prokaryotic diversity and its limits. S. 10494?10499.
Proceedings of the National Academy of Sciences
[online]. 2002-08-06 [cit. 2021-01-06]. Ro?. 99, ?is. 16, s. 10494?10499.
Dostupne online
.
DOI
10.1073/pnas.142680199
.
PMID
12097644
. (anglicky)
- ↑
SCHLOSS, Patrick D.; HANDELSMAN, Jo. Status of the Microbial Census. S. 686?691.
Microbiology and Molecular Biology Reviews
[online]. 2004-12 [cit. 2021-01-06]. Ro?. 68, ?is. 4, s. 686?691.
Dostupne online
.
DOI
10.1128/MMBR.68.4.686-691.2004
.
PMID
15590780
. (anglicky)
- ↑
WHITMAN, William B.; COLEMAN, David C.; WIEBE, William J. Prokaryotes: The unseen majority. S. 6578?6583.
Proceedings of the National Academy of Sciences
[online]. 1998-06-09 [cit. 2021-01-06]. Ro?. 95, ?is. 12, s. 6578?6583.
Dostupne online
.
DOI
10.1073/pnas.95.12.6578
.
PMID
9618454
. (anglicky)
- ↑
KLOTZ, Irene Mona.
Lichen love space
[online]. 2005.
Dostupne online
. (anglicky)
- ↑
a
b
c
d
ROSYPAL, Stanislav.
Bakteriologie a virologie
. Praha: Scientia, 1994.
- ↑
Moran, Laurence A.
Sandwalk: Multicellular Bacteria
[online]. 2008.
Dostupne online
.
- ↑
BASSLER, Bonnie L.; LOSICK. Bacterially speaking.
Cell
. 2006-04-21, ro?. 125, ?is. 2, s. 237?246.
Dostupne online
.
ISSN
0092-8674
.
DOI
10.1016/j.cell.2006.04.001
.
- ↑
FALCAO, Juliana P., Faith Sharp, Vanessa Sperandio. Cell-to-cell signaling in intestinal pathogens.
Current Issues in Intestinal Microbiology
. 2004-03, ro?. 5, ?is. 1.
Dostupne online
.
ISSN
1466-531X
.
- ↑
BASSLER, Bonnie.
The secret, social lives of bacteria
[online]. TEDtalksDirector (P?isp?vatel), 2009-04-08.
Dostupne online
.
- ↑
KYSILKA, Ji?i; KRMEN?IK, Pavel.
Toxicon ? Escherichia coli
[online].
Dostupne online
.
- ↑
ANGERT, Esther R.; CLEMENTS, Kendall D.; PACE, Norman R. The largest bacterium. S. 239?241.
Nature
[online]. 1993-03 [cit. 2021-01-06]. Ro?. 362, ?is. 6417, s. 239?241.
Dostupne online
.
DOI
10.1038/362239a0
. (anglicky)
- ↑
WUETHRICH, Bernice. Giant Sulfur-Eating Microbe Found. S. 415?415.
Science
[online]. 1999-04-16 [cit. 2021-01-06]. Ro?. 284, ?is. 5413, s. 415?415.
Dostupne online
.
ISSN
1095-9203
.
DOI
10.1126/science.284.5413.415
.
PMID
10232982
. (anglicky)
- ↑
PENNISI, Elizabeth. Largest bacterium ever discovered has an unexpectedly complex cell.
Science
[online]. American Association for the Advancement of Science, 2022-02-23 [cit. 2022-02-28].
Dostupne online
.
ISSN
1095-9203
.
DOI
10.1126/science.ada1620
. (anglicky)
- ↑
VOLLAND, Jean-Marie, et al. A centimeter-long bacterium with DNA compartmentalized in membrane-bound organelles.
BioRχiv
[online]. Cold Spring Harbor Laboratory, 2022-02-18 [cit. 2022-02-28]. Preprint.
Dostupne online
.
DOI
10.1101/2022.02.16.480423
. (anglicky)
- ↑
a
b
c
d
e
f
g
h
i
j
k
l
m
n
MURRAY, Patrick R.; ROSENTHAL, Ken S.; PFALLER, Michael A.
Medical Microbiology, Fifth edition
. [s.l.]: Elsevier, 2005. (anglicky)
- ↑
FOLK, Robert.
Nanobacteria: surely not figments, but what under heaven are they?
[online]. 1997.
Dostupne v archivu
po?izenem z
originalu
. (anglicky)
- ↑
HART, Stephen.
Small World
[online]. 2003.
Dostupne online
. (anglicky)
- ↑
PETR, Jaroslav.
Nanobakterie jsou ne?ive
[online]. Osel.cz, 2008.
Dostupne online
.
- ↑
a
b
c
d
e
f
CAMPBELL, N. A.; REECE, J. B.
Biologie
. [s.l.]: Computer press, 2006.
- ↑
?I?EK, Alois.
Praktika z veterinarni bakteriologie a mykologie
. Brno: VFU Brno, 1999.
- ↑
a
b
c
d
TODAR, Kenneth.
Textbook of Bacteriology
[online]. 2008.
Dostupne online
. (anglicky)
- ↑
?EPI?KA, Ivan, et al.
Pohyb
. Praha: Ust?edni komise biologicky olympiady, NIDM ?R, 2005.
Dostupne online
.
- ↑
a
b
CASJENS, Sherwood, Wai Mun Huang. Linear chromosomal physical and genetic map of
Borrelia burgdorferi
, the Lyme disease agent.
Molecular Microbiology
. 1993, ro?. 8, ?is. 5, s. 967?980.
Dostupne online
.
DOI
10.1111/j.1365-2958.1993.tb01641.x
.
- ↑
SLATER, Steven C.; GOLDMAN, Barry S.; GOODNER, Brad; SETUBAL, Joao C.; FARRAND, Stephen K.; NESTER, Eugene W.; BURR, Thomas J. Genome Sequences of Three Agrobacterium Biovars Help Elucidate the Evolution of Multichromosome Genomes in Bacteria. S. 2501?2511.
Journal of Bacteriology
[online]. 2009-04-15 [cit. 2021-01-06]. Ro?. 191, ?is. 8, s. 2501?2511.
Dostupne v archivu
po?izenem z
originalu
dne 2018-06-02.
DOI
10.1128/JB.01779-08
. (anglicky)
- ↑
FUERST, John A. Intracellular Compartmentation in Planctomycetes.
Annual Review of Microbiology
. 2005-05-23, ro?. 59, s. 299?328.
Dostupne online
.
DOI
10.1146/annurev.micro.59.030804.121258
.
Archivovano
30. 5. 2020 na
Wayback Machine
.
- ↑
LINDSAY, Margaret R., Richard I. Webb, Marc Strous, Mike S. Jetten, Margaret K. Butler, Rebecca J. Forde, John A. Fuerst. Cell compartmentalisation in planctomycetes: novel types of structural organisation for the bacterial cell.
Archives of Microbiology
. 2004-02-19, ro?. 175, ?is. 6, s. 413?429.
Dostupne online
.
DOI
10.1007/s002030100280
.
[
nedostupny zdroj
]
- ↑
SANTARELLA-MELLWIG, Rachel; FRANKE, Josef; JAEDICKE, Andreas, Matyas Gorjanacz, Ulrike Bauer, Aidan Budd, Iain W. Mattaj, Damien P. Devos. The Compartmentalized Bacteria of the Planctomycetes-Verrucomicrobia-Chlamydiae Superphylum Have Membrane Coat-Like Proteins. S. e1000281.
PLoS Biology
[online]. 19. leden 2010 [cit. 2010-01-21]. Svazek 8, ?is. 1, s. e1000281.
Dostupne online
. PDF
[1]
.
ISSN
1545-7885
.
DOI
10.1371/journal.pbio.1000281
. (anglicky)
- ↑
Introduction to the Archaean
[online]. University of California [cit. 2008-04-23].
Dostupne v archivu
po?izenem dne 2008-05-01. (anglicky)
- ↑
Looking for the last universal common ancestor of all living organisms.
phys.org
[online]. [cit. 2023-11-21].
Dostupne online
.
- ↑
SCHOPF, J. William. Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic. S. 6735?6742.
Proceedings of the National Academy of Sciences
[online]. 1994-07-19 [cit. 2021-01-06]. Ro?. 91, ?is. 15, s. 6735?6742.
Dostupne online
.
DOI
10.1073/pnas.91.15.6735
. (anglicky)
- ↑
DELONG, Edward F.; PACE, Norman R. Environmental Diversity of Bacteria and Archaea. S. 470?478.
Systematic Biology
[online]. 2001-08-01 [cit. 2021-01-06]. Ro?. 50, ?is. 4, s. 470?478.
Dostupne online
.
DOI
10.1080/10635150118513
. (anglicky)
- ↑
BROWN, James R.; DOOLITTLE, W. Ford. Archaea and the prokaryote-to-eukaryote transition.. S. 456?502.
Microbiology and Molecular Biology Reviews
[online]. 1997-12 [cit. 2021-01-06]. Ro?. 61, ?is. 4, s. 456?502.
Dostupne online
.
ISSN
1092-2172
.
PMID
9409149
. (anglicky)
- ↑
DI GIULIO, Massimo. The Universal Ancestor and the Ancestor of Bacteria Were Hyperthermophiles. S. 721?730.
Journal of Molecular Evolution
[online]. 2003-12 [cit. 2021-01-06]. Ro?. 57, ?is. 6, s. 721?730.
Dostupne online
.
DOI
10.1007/s00239-003-2522-6
. (anglicky)
- ↑
BATTISTUZZI, Fabia U; FEIJAO, Andreia; HEDGES, S Blair. A genomic timescale of prokaryote evolution: insights into the origin of methanogenesis, phototrophy, and the colonization of land. S. 44.
BMC Evolutionary Biology
[online]. 2004 [cit. 2021-01-06]. Ro?. 4, ?is. 1, s. 44.
Dostupne online
.
DOI
10.1186/1471-2148-4-44
.
PMID
15535883
. (anglicky)
- ↑
CAVALIER-SMITH, Thomas. Cell evolution and Earth history: stasis and revolution. S. 969?1006.
Philosophical Transactions of the Royal Society B: Biological Sciences
[online]. 2006-06-29 [cit. 2021-01-06]. Ro?. 361, ?is. 1470, s. 969?1006.
Dostupne online
.
DOI
10.1098/rstb.2006.1842
.
PMID
16754610
. (anglicky)
- ↑
CAVALIER-SMITH, Thomas. Rooting the tree of life by transition analyses. S. 19.
Biology Direct
[online]. 2006 [cit. 2021-01-06]. Ro?. 1, ?is. 1, s. 19.
Dostupne online
.
DOI
10.1186/1745-6150-1-19
.
PMID
16834776
. (anglicky)
- ↑
WOODS, Gail L.; WALKER, David H. Detection of infection or infectious agents by use of cytologic and histologic stains.. S. 382?404.
Clinical Microbiology Reviews
[online]. 1996-07-01 [cit. 2021-01-07]. Ro?. 9, ?is. 3, s. 382?404.
Dostupne online
.
DOI
10.1128/CMR.9.3.382
.
PMID
8809467
. (anglicky)
- ↑
LOUIE, Marie; LOUIE, Lisa; SIMOR, Andrew E. The role of DNA amplification technology in the diagnosis of infectious diseases. S. 301?309.
CMAJ
[online]. 2000-08-08 [cit. 2021-01-06]. Ro?. 163, ?is. 3, s. 301?309.
Dostupne online
.
ISSN
0820-3946
. (anglicky)
- ↑
Thomson R., Bertram H. Laboratory diagnosis of central nervous system infections.
Infect Dis Clin North Am
. 2001, ro?. 15, ?is. 4, s. 1047?71.
Dostupne online
. (anglicky)
- ↑
OLSEN, Gary J.; WOESE, Carl R.; OVERBEEK, Ross A. The winds of (evolutionary) change: Breathing new life into microbiology. S. MCS??P402-1293, 205047.
Journal of Bacteriology
[online]. 1996-03-01 [cit. 2021-01-06]. S. MCS??P402-1293, 205047.
Dostupne online
.
DOI
10.2172/205047
.
PMID
8282683
. (anglicky)
- ↑
OREN, Aharon; GARRITY, George M. Valid publication of the names of forty-two phyla of prokaryotes.
International Journal of Systematic and Evolutionary Microbiology
[online]. 2021-10-20 [cit. 2024-03-25]. Ro?. 71, ?is. 10.
DOI
10.1099/ijsem.0.005056
.
PMID
34694987
. (anglicky)
- ↑
OREN, Aharon; GOKER, Markus. Candidatus List. Lists of names of prokaryotic Candidatus phyla.
International Journal of Systematic and Evolutionary Microbiology
[online]. 2023-05-09. Ro?. 73, ?is. 5.
DOI
10.1099/ijsem.0.005821
.
PMID
37159402
. (anglicky)
- ↑
LPSN. Domain
Bacteria
.
Dostupne online
(anglicky)
- ↑
RUGGIERO, Michael A.; GORDON, Dennis P.; ORRELL, Thomas M., BAILLY, Nicolas; BOURGOIN, Thierry; BRUSCA, Richard C.;
CAVALIER-SMITH, Thomas
; GUIRY, Michael D.; KIRK, Paul M. A Higher Level Classification of All Living Organisms.
PLoS ONE
[online]. 29. duben 2015. Svazek 10, ?is. 4: e0119248.
Dostupne online
. PPT
[2]
. PNG
[3]
.
ISSN
1932-6203
.
DOI
10.1371/journal.pone.0130114
.
PMID
25923521
. (anglicky)
- ↑
RUGGIERO, Michael A.; GORDON, Dennis P.; ORRELL, Thomas M., BAILLY, Nicolas; BOURGOIN, Thierry; BRUSCA, Richard C.;
CAVALIER-SMITH, Thomas
; GUIRY, Michael D.; KIRK, Paul M. Correction: A Higher Level Classification of All Living Organisms.
PLoS ONE
[online]. 11. ?erven 2015. Svazek 10, ?is. 6: e0130114.
Dostupne online
. PPT
[4]
. PNG
[5]
.
ISSN
1932-6203
.
DOI
10.1371/journal.pone.0130114
.
PMID
26068874
. (anglicky)
- ↑
OREN, Aharon. Emendation of Principle 8, Rules 5b, 8, 15, 33a, and Appendix 7 of the International Code of Nomenclature of Prokaryotes to include the categories of kingdom and domain.
International Journal of Systematic and Evolutionary Microbiology
[online]. Microbiology Society, 2023-11-01 [cit. 2024-05-28]. Ro?. 73, ?is. 11.
ISSN
1466-5034
.
DOI
10.1099/ijsem.0.006123
.
PMID
37909283
. (anglicky)
- ↑
GOKER, Markus; OREN, Aharon. Valid publication of names of two domains and seven kingdoms of prokaryotes.
International Journal of Systematic and Evolutionary Microbiology
[online]. Microbiology Society, 2024-01-22 [cit. 2024-05-28]. Ro?. 74, ?is. 1.
DOI
10.1099/ijsem.0.006242
. (anglicky)
- ↑
a
b
c
KAPRALEK, Franti?ek.
Fyziologie bakterii
. [s.l.]: Statni pedagogicke nakladatelstvi, 1986. S. 603.
- ↑
ANGERT, Esther R. Alternatives to binary fission in bacteria. S. 214?224.
Nature Reviews Microbiology
[online]. 2005-03 [cit. 2021-01-07]. Ro?. 3, ?is. 3, s. 214?224.
Dostupne online
.
DOI
10.1038/nrmicro1096
. (anglicky)
- ↑
?IHOVA AMBRO?OVA, J.
Sinice, t?ida Cyanophyceae, odd?leni Cyanophyta
[online]. V?CHT Praha.
Dostupne online
.
- ↑
LINDQUIST, John.
Nutrition and Cultivation of Bacteria ? Page 1
[online]. University of Wisconsin [cit. 2008-05-15].
Dostupne v archivu
po?izenem dne 09-05-2008. (anglicky)
- ↑
Ustav biochemie a mikrobiologie V?CHT Praha.
Kultivace mikroorganism?
[online]. Vysoka ?kola chemicko-technologicka v Praze.
Dostupne online
.
[
nedostupny zdroj
]
? neplatny odkaz !
- ↑
HINNEBUSCH, Joe; TILLY, Kit. Linear plasmids and chromosomes in bacteria. S. 917?922.
Molecular Microbiology
[online]. 1993-12 [cit. 2021-01-06]. Ro?. 10, ?is. 5, s. 917?922.
Dostupne online
.
DOI
10.1111/j.1365-2958.1993.tb00963.x
.
PMID
7934868
. (anglicky)
- ↑
BENNETT, G. M.; MORAN, N. A. Small, smaller, smallest: the origins and evolution of ancient dual symbioses in a Phloem-feeding insect.
Genome Biol Evol
. 2013, ro?. 5, ?is. 9, s.
1675
?88.
Dostupne online
.
DOI
10.1093/gbe/evt118
.
PMID
23918810
.
- ↑
PRADELLA, Silke; HANS, Astrid; SPROER, Cathrin; REICHENBACH, Hans; GERTH, Klaus; BEYER, Stefan. Characterisation, genome size and genetic manipulation of the myxobacterium Sorangium cellulosum So ce56. S. 484?492.
Archives of Microbiology
[online]. 2002-12-01 [cit. 2021-01-06]. Ro?. 178, ?is. 6, s. 484?492.
Dostupne online
.
DOI
10.1007/s00203-002-0479-2
. (anglicky)
- ↑
BELFORT, M; REABAN, M E; COETZEE, T; DALGAARD, J Z. Prokaryotic introns and inteins: a panoply of form and function.. S. 3897?3903.
Journal of bacteriology
[online]. 1995 [cit. 2021-01-06]. Ro?. 177, ?is. 14, s. 3897?3903.
Dostupne v archivu
po?izenem z
originalu
dne 2020-12-01.
DOI
10.1128/jb.177.14.3897-3903.1995
. (anglicky)
- ↑
NEALSON, K. Post-Viking microbiology: new approaches, new data, new insights.
Orig Life Evol Biosph
. 1999, ro?. 29, ?is. 1, s. 73?93. (anglicky)
- ↑
XU, Jianping. Microbial ecology in the age of genomics and metagenomics: concepts, tools, and recent advances. S. 1713?1731.
Molecular Ecology
[online]. 2006 [cit. 2021-01-07]. Ro?. 15, ?is. 7, s. 1713?1731.
Dostupne online
.
DOI
10.1111/j.1365-294X.2006.02882.x
. (anglicky)
- ↑
a
b
c
ROSYPAL, Stanislav
.
Novy p?ehled biologie
. [s.l.]: Scientia, 2003. S. 797.
- ↑
ZILLIG, Wolfram. Comparative biochemistry of Archaea and Bacteria. S. 544?551.
Current Opinion in Genetics & Development
[online]. 1991-12 [cit. 2021-01-07]. Ro?. 1, ?is. 4, s. 544?551.
Dostupne online
.
DOI
10.1016/S0959-437X(05)80206-0
. (anglicky)
- ↑
HELLINGWERF, K. J.; CRIELAARD, W.; HOFF, W. D.; MATTHIJS, H. C. P.; MUR, L. R.; VAN ROTTERDAM, B. J. Photobiology of Bacteria. S. 331?347.
Antonie van Leeuwenhoek
[online]. 1994-12 [cit. 2021-01-06]. Ro?. 65, ?is. 4, s. 331?347.
Dostupne online
.
DOI
10.1007/BF00872217
.
PMID
7832590
. (anglicky)
- ↑
BRYANT, Donald A.; COSTAS, Amaya M. Garcia; MARESCA, Julia A.; CHEW, Aline Gomez Maqueo; KLATT, Christian G.; BATESON, Mary M.; TALLON, Luke J. Candidatus Chloracidobacterium thermophilum: An Aerobic Phototrophic Acidobacterium. S. 523?526.
Science
[online]. 2007-07-27 [cit. 2021-01-06]. Ro?. 317, ?is. 5837, s. 523?526.
Dostupne online
.
DOI
10.1126/science.1143236
. (anglicky)
- ↑
University of California ? Museum of Paleontology.
Bacteria: Life History and Ecology
[online]. [cit. 2008-04-25].
Dostupne v archivu
po?izenem dne 2008-05-08. (anglicky)
- ↑
POOLE, Anthony M.; PENNY, David. Evaluating hypotheses for the origin of eukaryotes. S. 74?84.
BioEssays
[online]. 2007-01 [cit. 2021-01-05]. Ro?. 29, ?is. 1, s. 74?84.
Dostupne online
.
DOI
10.1002/bies.20516
.
PMID
17187354
. (anglicky)
- ↑
DYALL, S. D. Ancient Invasions: From Endosymbionts to Organelles. S. 253?257.
Science
[online]. 2004-04-09 [cit. 2021-01-05]. Ro?. 304, ?is. 5668, s. 253?257.
Dostupne v archivu
po?izenem z
originalu
dne 2021-08-30.
DOI
10.1126/science.1094884
. (anglicky)
- ↑
LANG, B. Franz; GRAY, Michael W.; BURGER, Gertraud. Mitochondrial Genome Evolution and the Origin of Eukaryotes. S. 351?397.
Annual Review of Genetics
[online]. 1999-12 [cit. 2021-01-05]. Ro?. 33, ?is. 1, s. 351?397.
Dostupne v archivu
po?izenem z
originalu
dne 2021-03-30.
DOI
10.1146/annurev.genet.33.1.351
. (anglicky)
- ↑
MCFADDEN, Geoffrey Ian. Endosymbiosis and evolution of the plant cell. S. 513?519.
Current Opinion in Plant Biology
[online]. 1999-12 [cit. 2021-01-06]. Ro?. 2, ?is. 6, s. 513?519.
Dostupne online
.
DOI
10.1016/S1369-5266(99)00025-4
. (anglicky)
- ↑
a
b
?EPI?KA, Ivan; KOLA?, Filip; SYNEK, Petr.
Mutualismus, vzajemn? prosp??na symbioza; P?ipravny text ? biologicka olympiada 2007?2008
. Praha: NIDM ?R, 2007. S. 87.
- ↑
RIGOTTIER-GOIS, Lionel; ROCHET, Violaine; GARREC, Nathalie; SUAU, Antonia; DORE, Joel. Enumeration of Bacteroides Species in Human Faeces by Fluorescent in situ Hybridisation Combined with Flow Cytometry Using 16S rRNA Probes. S. 110?118.
Systematic and Applied Microbiology
[online]. 2003-01 [cit. 2021-01-06]. Ro?. 26, ?is. 1, s. 110?118.
Dostupne online
.
DOI
10.1078/072320203322337399
.
PMID
12747418
. (anglicky)
- ↑
SENDER, Ron; FUCHS, Shai; MILO, Ron. Are We Really Vastly Outnumbered? Revisiting the Ratio of Bacterial to Host Cells in Humans.
Cell
. 2016-01-28, ro?. 164, ?is. 3, s. 337?340.
PMID
26824647
.
Dostupne online
[cit. 2016-02-12].
ISSN
1097-4172
.
DOI
10.1016/j.cell.2016.01.013
.
PMID
26824647
.
- ↑
GUARNER, Francisco; MALAGELADA, Juan-R. Gut flora in health and disease.
Lancet (London, England)
. 2003-02-08, ro?. 361, ?is. 9356, s. 512?519.
PMID
12583961
.
Dostupne online
[cit. 2016-02-12].
ISSN
0140-6736
.
DOI
10.1016/S0140-6736(03)12489-0
.
PMID
12583961
.
- ↑
MUELLER, C; MACPHERSON, A. J. Layers of mutualism with commensal bacteria protect us from intestinal inflammation.
Gut
. Unor 2006, ro?. 55(2), s. 276?284.
Dostupne online
. (anglicky)
- ↑
Anonym.
Microbial Flora of Skin
[online]. [cit. 2008-04-24].
Dostupne v archivu
po?izenem dne 25-02-2008. (anglicky)
? neplatny odkaz !
- ↑
FEREN?IK, M.; ROVENSKY, J.; SHOENFELD, Y; MA?HA, V.
Imunitni system; informace pro ka?deho
. 1. ?eske. vyd. Praha: Grada Publishing, 2005.
- ↑
?lov?k jako ekosystem.
ekonom.cz
[online]. 2012-10-11 [cit. 2021-01-05].
Dostupne online
.
- ↑
http://www.jhnewsandguide.com/jackson_hole_daily/our_world_people/our_world/unprecedented-germ-diversity-found-in-remote-amazonian-tribe/article_baa6f54a-5722-5ea2-a014-462f5bcd1ff6.html
[
nedostupny zdroj
]
? Unprecedented germ diversity found in remote Amazonian tribe ? neplatny odkaz !
- ↑
JOHNSON, M.E.; LUCEY, J.A. Major Technological Advances and Trends in Cheese. S. 1174?1178.
Journal of Dairy Science
[online]. 2006-04 [cit. 2021-01-07]. Ro?. 89, ?is. 4, s. 1174?1178.
Dostupne online
.
DOI
10.3168/jds.S0022-0302(06)72186-5
.
PMID
16537950
. (anglicky)
- ↑
HAGEDORN, Scott; KAPHAMMER, Bryan. Microbial Biocatalysis in the Generation of Flavor and Fragrance Chemicals. S. 773?800.
Annual Review of Microbiology
[online]. 1994-10 [cit. 2021-01-06]. Ro?. 48, ?is. 1, s. 773?800.
Dostupne v archivu
po?izenem z
originalu
dne 2021-03-08.
DOI
10.1146/annurev.mi.48.100194.004013
. (anglicky)
- ↑
K?DELA, Vaclav; NOVACKY, Anton; FUCIKOVSKY, Leopold.
Rostlinoleka?ska bakteriologie
. [s.l.]: Academia, 2002. S. 346.
- ↑
BALL, Christopher.
Genetics and Breeding of Industrial Microorganisms
. [s.l.]: CRC Press, 1984.
Dostupne online
.
ISBN
0849356725
. S. 203 ♀4 url =
http://books.google.com/books?id=3wiwjhMN5DQC&printsec=frontcover&dq=Mutation+Breeding+bacteria&as_brr=3&hl=cs&source=gbs_summary_r&cad=0
.
- ↑
Beychok, Milton R. Performance on surface-aerated basins.
Chemical Engineering Progress, Symposium series
. 1971, ro?. 67, ?is. 107, s. 322?339.
Dostupne v archivu
po?izenem dne 14-11-2007. (anglicky)
Archivovano
14. 11. 2007 na
Wayback Machine
. ? neplatny odkaz !
- ↑
Septic Systems for Waste Water Disposal, on-line version of American Ground Water Trust's
Consumer Awareness Information Pamphlet
[online]. [cit. 2008-05-20].
Dostupne v archivu
po?izenem dne 17-05-2008. (anglicky)
? neplatny odkaz !
- ↑
COHEN, Yehuda. Bioremediation of oil by marine microbial mats. S. 189?193.
International Microbiology
[online]. 2002-12-01 [cit. 2021-01-06]. Ro?. 5, ?is. 4, s. 189?193.
Dostupne online
.
DOI
10.1007/s10123-002-0089-5
.
PMID
12497184
. (anglicky)
- ↑
LIESE, Andreas; VILLELA FILHO, Murillo. Production of fine chemicals using biocatalysis. S. 595?603.
Current Opinion in Biotechnology
[online]. 1999-12-01 [cit. 2021-01-06]. Ro?. 10, ?is. 6, s. 595?603.
Dostupne online
.
DOI
10.1016/S0958-1669(99)00040-3
. (anglicky)
- ↑
ARONSON, Arthur I.; SHAI, Yechiel. Why Bacillus thuringiensis insecticidal toxins are so effective: unique features of their mode of action. S. 1?8.
FEMS Microbiology Letters
[online]. 2001-02 [cit. 2021-01-06]. Ro?. 195, ?is. 1, s. 1?8.
Dostupne online
.
DOI
10.1111/j.1574-6968.2001.tb10489.x
.
PMID
11166987
. (anglicky)
- ↑
WALSH, Gary. Therapeutic insulins and their large-scale manufacture. S. 151?159.
Applied Microbiology and Biotechnology
[online]. 2005-04 [cit. 2021-01-06]. Ro?. 67, ?is. 2, s. 151?159.
Dostupne online
.
DOI
10.1007/s00253-004-1809-x
.
PMID
15580495
. (anglicky)
- ↑
GRAUMANN, Klaus; PREMSTALLER, Andreas. Manufacturing of recombinant therapeutic proteins in microbial systems. S. 164?186.
Biotechnology Journal
[online]. 2006-02 [cit. 2021-01-06]. Ro?. 1, ?is. 2, s. 164?186.
Dostupne online
.
DOI
10.1002/biot.200500051
.
PMID
16892246
. (anglicky)
- ↑
SERRES, Margrethe H; GOPAL, Shuba; NAHUM, Laila A; LIANG, Ping; GAASTERLAND, Terry; RILEY, Monica. A functional update of the Escherichia coli K-12 genome. S. research0035.1.
Genome Biology
[online]. 2001 [cit. 2021-01-06]. Ro?. 2, ?is. 9, s. research0035.1.
Dostupne online
.
DOI
10.1186/gb-2001-2-9-research0035
.
PMID
11574054
. (anglicky)
- ↑
ALMAAS, E.; KOVACS, B.; VICSEK, T.; OLTVAI, Z. N.; BARABASI, A.-L. Global organization of metabolic fluxes in the bacterium Escherichia coli. S. 839?843.
Nature
[online]. 2004-02 [cit. 2021-01-06]. Ro?. 427, ?is. 6977, s. 839?843.
Dostupne online
.
DOI
10.1038/nature02289
. (anglicky)
- ↑
HIEI, Yukoh; KOMARI, Toshihiko. Agrobacterium-mediated transformation of rice using immature embryos or calli induced from mature seed. S. 824?834.
Nature Protocols
[online]. 2008-05 [cit. 2021-01-06]. Ro?. 3, ?is. 5, s. 824?834.
Dostupne online
.
DOI
10.1038/nprot.2008.46
.
PMID
18451790
. (anglicky)
- ↑
PRESCOTT, Lansing M.
Microbiology
. 5. vyd. Boston: McGraw-Hill
Dostupne online
.
ISBN
0-07-232041-9
,
ISBN
978-0-07-232041-1
.
OCLC
46319741
S. 326.
- ↑
PETR, Jaroslav. CRISPR: p?esna st?elba na geneticke cile.
Vesmir
. 2015.
Dostupne online
.
- BEDNA?, Marek, et al.
Leka?ska mikrobiologie : bakteriologie, virologie, parazitologie
. Praha: Triton, 1996. 560 s.
ISBN
80-2380-297-6
.
- VOTAVA, Miroslav.
Leka?ska mikrobiologie obecna
. Brno: Neptun, 2001. 247 s.
ISBN
80-902896-2-2
.
- VOTAVA, Miroslav.
Leka?ska mikrobiologie specialni
. Brno: Neptun, 2003. 495 s.
ISBN
80-902896-6-5
.
- KLABAN, Vladimir.
Sv?t mikrob?; ilustrovany lexikon mikrobiologie ?ivotniho prost?edi
. Hradec Kralove: Gaudeamus, 2001. 416 s.
ISBN
80-7041-687-4
.
- KAPRALEK, Franti?ek.
Zaklady bakteriologie
. Praha: Karolinum, 2000. 241 s.
ISBN
80-7184-811-5
.
- KAPRALEK, Franti?ek.
Fyziologie bakterii
. [s.l.]: Statni pedagogicke nakladatelstvi, 1986. 603 s.
- LHOTSKY, Josef.
Uvod do studia symbiotickych interakci mikroorganism?. Novy pohled na viry a bakterie
. Praha, Academia, 2015, 208 s.
- K?DELA, Vaclav; NOVACKY, Anton; FUCIKOVSKY, Leopold.
Rostlinoleka?ska bakteriologie
. Praha: Academia, 2002. 346 s.
ISBN
80-200-0899-3
.
- ROSYPAL, Stanislav
.
Bakteriologie a virologie
. Praha: Scientia, 1994. 67 s.
ISBN
80-85827-16-6
.
- ROSYPAL, Stanislav
.
Novy p?ehled biologie
. Praha: Scientia, 2003. 797 s.
ISBN
80-7183-268-5
.