 Mapa svijeta prema boji ko?e autohtonog
stanovni?tva
Mapa svijeta prema boji ko?e autohtonog
stanovni?tva
Rasa
, u
op?ebiolo?kom
smislu, je
taksonomska
kategorija između (ni?eg)
varijeteta
(
varietas
) i (hijerarhijski vi?e)
podvrste
(
subspecies
). U opisu infraspekcijske promjenljivosti, to je kategorija
zoolo?ke
nomenklature, dok u
botanici
odgovaraju?i rang ima
sorta
.
[1]
[2]
[3]
Rasa je ustvari dru?tveni koncept koji se primjenjuje za kategorizaciju ljudi u velike i razli?ite populacije ili grupe po
anatomskoj
,
kulturnoj
,
etni?koj
,
geneti?kij
,
geografskoj
,
historijskoj
,
jezi?koj
,
vjerskoj
, i / ili
socijalnoj
osobenosti. Ovaj termin se najprije koristio za govornike zajedni?kog
jezika
, a zatim da ozna?i nacionalnu pripadnosti. U17. stolje?u, ljudi su po?eli koristiti termin u smislu diferencijacije po vidljivim
fizi?kim
osobinama
. Takva upotreba promovirane hijerarhije bila je prikladnija za razli?ite etni?ke grupe. Po?ev od 19. stolje?a, izraz se ?esto koristi u taksonomskom smislu za ozna?avanje
geneti?ke
diferencijacije ljudskih populacija na bazi
fenotipskih
svojstava.
Socijalne koncepcije i grupiranje rasa varirale su tokom vremena, uklju?uju?i ?folk taksonomiju“ koja definira ?esencijalne tipove“ pojedinaca na osnovu percepcije njihovih
osobina
. Biolo?ki
esencijalizam
nanstvenici smatraju zastarjelm i op?enito obeshrabruju rasna obja?njenja za kolektivnu diferencijaciju po
fizi?kim
i
etolo?kim
osobinama
.
Iako postoji ?iroka suglasnost nau?ne zajednice da su
esencijalisti?ki
i tipolo?ki koncepti rase neodr?ivi, nau?nici ?irom svijeta i dalje nastavljaju takvu utrku na ?irokoj lepezi razli?itih na?ina, od kojih neke imaju i esencijalisti?ke implikacije. Dok neki istra?iva?i ponekad koriste koncept rase da opi?u razlike između nejasnih skupova osobina, drugi ukazuju na to da se ideja rase ?esto koristi u naivnom ili pojednostavljenom zna?enju pa tvrde u svekolikoj
ljudskoj
populaciji rasa nema taksonomski zna?aj, ukazuju?i da svi ljudi dana?njice pripadaju istoj
vrsti
(
Homo sapiens
) i podvrsti
Homo sapiens sapiens
.
Od druge polovine 20. stolje?a, razmimoila?enja u ideologijama i teorijama koje su se zasnivale na
bioantropolo?kim
i
fiziolo?kim
istra?ivanjima 19. stolje?a dovela je do takmi?enja u upotrebi rije?i, koje samo po sebi postaje problemati?no. Iako se jo? uvijek koristi u mnogim kontekstima, rasa se ?esto zamijenjuje drugim rije?ima koje su manje dvosmislene i nisu nabijene emocijama, kao ?to su
populacija
,
ljudi
etni?ke grupe
ili zajednice, ovisno o kontekstu.
Prosje?na genetska udaljenost Fst između ljudskih populacija iznosi otprilike 0,125. Lewontin (1972.) tvrdi da to predstavlja malu rasnu varijaciju.
[4]
Harpending (2002.), s druge strane, tvrdi da takva udaljenost implicira srodstvo između jedinki iste populacije ekvivalentno srodstvu između polubra?e u populaciji koja se parenjem odvija nasumi?no, te da je osoba određene populacije genetski bli?a nepovezanoj osobi iste populacije nego polubra?i mije?ane rase.
[5]
Ljudske rase
je termin koji se koristi da bi se pojam
stanovni?tvo
(
populacija
) razlikovao od pojma
?ovjek
, iako se biolo?ki pojam
rasa
ne odnosi isklju?ivo na populacijsku bioraznolikost
Homo sapiensa
. Prema "klasi?nom" konceptu "rase", kao najuobi?ajenija rasna obilje?ja su uzimanane najuo?ljivije
morfolo?ke
osobine
, kao ?to su
boja ko?e
,
boja o?iju
,
boja kose
i
forma kose
, te fizionomija
lica
,
stas
,
tjelesna konstitucija
i druga markantna svojstva, koja su u promatranoj populaciji uo?ljivo frekventnija nego u ostalima.
[6]
[7]
[8]
[9]
[10]
Savremeni koncept "rase" po?iva na komparativnim analizama
geneti?ke
strukture
populacije
i "međurasne razlike" posmatra kao i sve ostale međupopulacijske.
[11]
[12]
[13]
[14]
Rasne koncepcije, kao i kategorizacija specifi?nih rasnih grupacija i njihovih grana i ogranaka, oduvijek variraju, zavisno o
kulturi
i epohi kojoj pripadaju i suvi?e ?esto su bile kontroverzne prirode (zbog njihovog uticaja na socijalni
identitet
i
politiku
odnosa prema tom identitetu.
Grupna
biolo?ka
promjenljivost recentnog ?ovjeka na nivou "rasa" permanentno zaokuplja posebnu pozornost kako u oblasti
bioantropologije
,
genetike
,
evolucije
i
medicine
tako i
sociologije
,
filozofije
,
prava
,
historije
i ostalih humanisti?kih
nauka
pa i u naj?irem krugu obi?nih radoznalaca. Unato? tome, jo? uvijek nema usagla?enog nau?nog stava o kriterijima, smislu i prirodi "rasne" diferencijacije savremenog
?ovjeka
. Tako svjetski autoriteti u razli?itim oblastima bioantropologije (posebno humane genetike i evolucije) ocjenjuju da ni u jednoj drugoj oblasti
biologije
?ovjeka
do sada nije bilo toliko lutanja, "zbrke i nerazumijevanja".
[15]
To je, u velikoj mjeri, kauzalna posljedica minulih "rasnih" predrasuda i njima izazvanih reakcija. U ogromnoj su disproporciji stupanj op?eg nau?nog interesovanja za “rasnu” diferencijaciju ?ovje?anstva i nivo pouzdanosti prikupljenih egzaktnih podataka o
adaptacijskom
zna?aju ?ak i onih "rasnih" obilje?ja ?ija se varijacija naj?e??e uvr?tava u klju?ne kriterije kategorizacije u ovoj oblasti. Argumentirano se mo?e tvrditi da predstave o njihovoj adaptivnosti uglavnom po?ivaju tek na prihvatljivim nagađanjima i pretpostavkama.
[1]
[2]
[3]
[16]
Savremena bioantropolo?ka definicija pod "rasom" podrazumijeva jednu i/ili sistem srodnih ljudskih populacija osobene geneti?ke strukture, koja se prepoznatljivo razlikuje od one u drugim ? istovjetno definiranim ? ljudskim skupinama.
Sociobiolo?ko
jedinstvo evolucije
?ovjeka
je uvjetovalo jednu od krucijalnih specifi?nosti ljudskih "rasa" ? za razliku od animalnih, one nisu tranzicijske jedinice procesa nastajanja novih
vrsta
(
specijacije
).
Antropogeni
faktori su, naime, u tolikoj mjeri transformirali op?ebiolo?ku ulogu i funkciju
geneti?ke
varijabilnosti da je
kultura
postala najmo?nija forma prilagođavanja ljudske vrste ?ivotnoj sredini. Ta ?injenica nije kontradiktorna evidencijama da su okviri i potencijali i ovog oblika adaptacije uveliko određeni op?im geneti?kim, odnosno bioantropolo?kim pojavama i procesima. Dostignu?a civilizacije, naime, favorizuju bogatstvo mogu?nosti i formi osiguranja ?ivotne egzistencije i time pove?avaju ?ansu razli?itih
genotipova
u formiranju genskog fonda narednih
generacija
. Na taj na?in, kulturni progres je istovremeno omogu?io i biolo?ko jedinstvo ljudske vrste i izrazitu varijaciju i
polimorfizam
. U ovom integralnom kompleksu progresivno se kumulirala i njena posebno bitna osobenost ?
specijalizacija
za despecijalizaciju
.
Najuo?ljivija "rasna" obilje?ja, ustvari, predstavljaju [adaptacija|[adaptacijske]] "uspomene" iz prethodnih etapa
hominizacije
, u kojima su djelovali op?i prirodni faktori evolucije, a posljedica su
selektivne
prilagodbe na razli?ite komplekse uticaja ?ivotne sredine. Budu?i da egzistencijalni prirodni resursi nisu homogeno distribuirani po ?itavoj Planeti, geneti?ki sastav razli?itih
populacija
(polimorfne i
kosmopolitske
)
ljudske
vrste
adaptirao se na veoma heterogene ?ivotne uvjete. Tako se argumentirano mo?e zaklju?iti da su najuo?ljivije spoljne “rasne” osobenosti (kao ?to su boja ko?e, boja kose i forma dlake, stas, konstitucija i op?i habitus, oblik lica i lubanje, oblik i boja o?iju i druga) imale ili imaju nagla?en adaptivni zna?aj. Njihove odgovaraju?e varijante i cjeline bile su selektivno sna?no podr?avane u razli?itim uvjetima insolacije, temperature, vlage, ishrane i niza drugih ?inilaca.
Boja ko?e
je najupadljivija superficijelna "rasna" osobenost, koja je, obi?no, uva?avana kao jedan od najzna?ajnijih (a u klasi?nim pristupima i odlu?uju?ih) pokazatelja distinkcije "rasa". Poznavanje njenog funkcionalnog zna?aja u op?em koadaptivnom
sistemu
?ovjeka
, međutim, daleko je ispod te razine.
Prostorno
?
geografska
distribucija
slabe i jake pigmentacije ljudskih skupina argumentira pretpostavku da tamna put predstavlja prilagodbu na intenzivnu insolaciju i vedro
nebo
, a da svijetla ima adaptivnu prednost u oblastima prohladne
klime
sa mnogo
magle
i obla?nih dana.
Takvu hipotezu podr?ava i
Glogerovo pravilo
, prema kojem su ?ivotinjske jedinke iz populacija tropskih vla?nih oblasti obi?no intenzivnije
pigmentirane
nego one iz hladnih i suhljih dijelova
areala
istih i srodnih
vrsta
. Najtamnoputiji savremeni ljudi ?ive između
Sahare
i
afri?kih
tropskih ki?nih
?uma
, a izrazito su pigmentirani i stanovnici
Melanezije
,
Nove Gvineje
, ju?ne
Indije
i autohtoni
Australci
. Centri radijacije leukodermije ili blondizma su u
sjevernoj
Evropi
, koju karakteziraju
klimati
sa malo sun?anih dana, posebno u onim podru?jima koja su bila zahva?ena
pleistocenskim
glacijacijama
(oledbama).
Mongolske
populacije se, međutim, ne uklapaju u ovu op?u sliku;
ameri?ki
Indijanci
(sup)tropskih krajeva su neznatno tamniji od svojih srodnika iz umjerenih i hladnih predjela, a sibirski palemongoloidi i
Eskimi
(ameri?kog
Arktika
) imaju ne?to vi?e
pigmenta
nego ?to bi bilo primjereno njihovom podneblju. Jedno od prihvatljivih obja?njenja tog fenomena moglo bi biti u vezi sa klimatskim prilikama arkti?ke oblasti (posebno u
Aziji
), gdje je prete?no suho i rijetko obla?no, sa blje?te?om
insolacijom
.
?esto se postavlja i pitanje funkcionalnog paralelizma u adaptivnoj ulozi preplanulosti svjetloputih i trajne ("rasne") tamnoputosti. Kao ?to je poznato, insolacijski inducirana pigmentacija kod osoba svijetle puti ima za?titnu ulogu u prevenciji bolnih opekotina.
UV
dio
spektra
(
talasne du?ine
oko 3.000
angstrema
) ozljeđuje ?ive povr?inske slojeve
ko?e
(posebno dermalni i
Malpigijev
), a preplanulost inducira za?titno deponovanje pigmenta u ?elijama poko?ice i zadebljavanje njenog ro?nog sloja (nakupljanjem ostataka mrtvih Malpigijevih
?elija
. Ro?ni sloj
apsorbira
UV
zrake
i spre?ava njihov prodor u dublje ? ?ive
?elije
, pri ?emu je efikasnost te za?titne funkcije u direktnoj srazmjeri sa stupnjem
pigmentiranosti
. Takva uloga inducirane pigmentacije je, dakle,
idioadaptacijske
prirode
, dok trajna (intenzivna) pigmentiranost ima
evolutivno
?
adaptivno
zna?enje.
Relativno ve?a adaptivna vrijednost svjetlijih nijansi ko?e u podnebljima sa malo
sun?anih
dana
također nije pouzdano potvrđena, a pretpostavlja se da bi mogla biti u vezi sa stimulacijom sinteze anti
rahiti?nog
vitamina
D u ?ivim ?elijama (na taj na?in ?to upija UV zrake). Tamna put odbija svega oko 10?20% vidljivog dijela
spektra
, tj. zna?ajno manje nego svijetla (40?50%), ?to pove?ava toplotno optere?enje. Taj porast podlo?nosti opasnostima od toplotnog udara, u podno?ljivim granicama, mo?e se i potpuno kompenzirati znojenjem na relativno ni?oj spoljnoj
temperaturi
od normalne. Također se vjeruje da je adaptivni zna?aj svijetle puti, a naro?ito njene varijante sa plavom
kosom
i plavim
o?ima
, mogao biti selektivno favorizovan tokom posljednje interglacijacije i nakon zadnje
glacijacije
(oledbe).
Postoje argumenti koji podr?avaju i pretpostavku da je pove?ana
pigmentiranost
također bila favorizovana i u vezi sa rakom ko?e. On je ?e??i među svjetloputim osobama, a u istoj kategoriji
pigmentacije
(i kod istih osoba) ?e??e se javlja na dijelovima ko?e koji su neposredno eksponirani suncu. S druge strane, smanjena osjetljivost osoba svijetle puti na nazebe i upale
zglobova
mogla je biti faktor pove?anja adaptivne vrijednosti ovog
fenotipa
u hladnijim podnebljima.
Smatra se da je ?esto pominjana
hipoteza
o “za?titnoj obojenosti” kao adaptaciji tamnoputih
lova?kih
plemena
na uvjete sjenovitih
?uma
? na razini neutemeljenih spekulacija.
Forma kose
, po svemu sude?i, i do recentnog doba je odr?ala svoj primarni adaptivni zna?aj; vunasta, spiralna, ku?trava i kovrd?ava kosa pripadnika tropskih populacija efikasnija su za?tita mozga od vrele insolacije nego ?to su, naprimjer, prava i valovita.
Tjelesna konstitucija
i oblik
tijela
također mogu imati nagla?en adaptivni zna?aj. Posmatranjem globalne distribucije tipova
konstitucije
, dimenzija i
oblika
tijela kod
?ivotinja
je zapa?eno nekoliko op?ih karakteristika (
osobina
) njihove
prostorne
(spacijalne) varijacije.
Bergmanovo pravilo
, naprimjer, po?iva na uo?enoj pojavi da su jedinke vrsta koje ?ive u hladnijim
klimatima
obi?no (prosje?no) krupnije od svojih srodnika iz toplijih krajeva.
Alenovo pravilo
se, pak, odnosi na pojavu da su istureni dijelovi tijela (
udovi
,
u?i
,
rep
i dr.) u hladnijim dijelovima areala iste ili srodnih
vrsta
obi?no kra?i (i manjih ostalih parametara) u odnosu na op?e tjelesne dimenzije. Obje ove pojave su u funkciji ?uvanja
tjelesne
toplote. U vezi s tim treba se prisjetiti da u proporcionalnom razvoju
organizma
povr?ina
raste po kvadratnoj, a
zapremina
i
masa
po kubnoj potenciji linearnih
mjera
. Manja povr?ina, dakle, odaje relativno manje
toplote
po jedinici mase, pa selektivnu prednost u razli?itim
podnebljima
imaju kontrastno druga?iji
fenotipovi
.
Karakteristi?ni kvantitativni odnos
tjelesne
visine
i mase u pojedinim dijelovima sveukupnog svjetskog
stanovni?tva
daje veoma indikativne podatke o mogu?im vezama između klimatskih prilika i tjelesne konstitucije (vidi prilo?enu tabelu!). Neki primjeri najvi?eg (Vatutsi ? 176 cm) i najni?eg (
Pigmeji
? 142 cm) rasta u ljudskoj populaciji pokazuju da stanovni?tvo vrelih i otvorenih predjela spada u kategoriju najve?ih iznosa tog koeficijenta, iako je njihova adaptacija i?la u dva razli?ita pravca; oni naginju ka visokom i suhonjavom stasu ili, jo? ?e??e, niskom stasu i vitkoj konstituciji (srazmjerno malog tijela i izdu?enih udova). Autohtoni stanovnici hladnih oblasti su naj?e??e korpulentni ? ?iroki, krupni i te?ki, odnosno zdepasti i de?mekasti, a skloni su i nagomilavanju potko?nog masnog tkiva (koje je veoma efikasan toplotni
izolator
). Nasuprot tome, salo u tropskim krajevima pove?ava opasnost od toplotnog iznurivanja. ?injenica da odje?a savremenog
?ovjeka
odr?ava određenu mikroklimu i na veoma niskim
temperaturama
ne zna?i da na individualnu
adaptabilnost
ne uti?e i tjelesna konstitucija (posebno u ekstremnim uvjetima). Osobito ne treba zanemariti ni ?injenicu da je adaptivnost
evolutivno
dinami?na kategorija. Također se pretpostavlja da termoizolaciona svojstva potko?ne masno?e obja?njavaju i karakteristi?nu strukturu
mongoloidnog
lica.
U hladnim oblastima
tijelo
je za?ti?eno
odje?om
, a
lice
mo?e biti eksponirano izrazito niskim temperaturama. Pripadnici ve?ine populacija iz centra radijacije mongoloida ?ivi u
klimatima
niskih
temperatura
, o?trih
mrazeva
i jakih vjetrova, pa njihova spljo?tena i dobro popunjena lica imaju manju relativnu povr?inu (na jedinicu mase); otpornija su na ozebline nego mr?ava lica istaknutih crta. Adaptivni zna?aj takvih svojstava potvrđuje i primjer skandinavskih Laponaca (
evropeidnog
porijekla) koji su
konvergencijom
(u sli?nim uvjetima ?ivotne sredine) stekli mnoga mongoloidna obilje?ja.
| Stanovni?tvo
|
Visina
(cm) -
h
|
Masa
(kg) -
m
|
h
/
m
|
| Kavkazoidi
|
| Finska
|
171,0
|
70,0
|
2,44
|
| SAD
|
173,9
|
70,2
|
2,48
|
| Island
|
173,6
|
66,1
|
2,55
|
| Engleska
|
166,3
|
64,5
|
2,57
|
| Francuska
|
172,5
|
67,0
|
2,57
|
| SAD
|
173,9
|
70,2
|
2,48
|
| Sicilija
|
169,1
|
65,0
|
2,65
|
| Maroko
|
168,9
|
63,8
|
2,65
|
| ?kotska
|
170,4
|
61,8
|
2,76
|
| Tunis
|
173,4
|
62,3
|
2,78
|
| Berberi
|
169,8
|
59,5
|
2,85
|
| Indija
: Marata
|
163,8
|
55,7
|
2,94
|
| Indija
: Benga
|
165,8
|
52,7
|
3,15
|
| Negroidi
|
| Jambasa
|
169,0
|
62,0
|
2,78
|
| Kirdi
|
166,5
|
57,3
|
2,90
|
| Baja
|
163,0
|
53,9
|
3,02
|
| Vatutsi
|
176,0
|
57,0
|
3,09
|
| Kikuju
|
164,5
|
51,9
|
3,56
|
| Pigmeji
|
142,2
|
39,9
|
3,56
|
| Efeji
|
143,8
|
39,8
|
3,61
|
| Bu?mani
|
155,8
|
40,4
|
3,86
|
| Mongoloidi
|
| Turkmenistan
: Kazasi
|
163,1
|
69,7
|
2,34
|
| Eskimi
|
161,2
|
62,9
|
2,56
|
| Sjeverna Koreja
|
168,0
|
61,0
|
2,75
|
| Ju?na Koreja
|
161,1
|
55,5
|
2,90
|
| Centralna
Kina
|
163,0
|
54,7
|
2,98
|
| Japan
|
160,9
|
53,0
|
3,04
|
| Sundan
|
159,8
|
51,9
|
3,08
|
| Anamiti
|
158,7
|
51,3
|
3,09
|
| Hong Kong
|
166,2
|
52,2
|
3,18
|
Fiziolo?ka adaptacija
je jedna od nezaobilaznih komponenti sistema odr?avanja
homeostaze
ljudskog
organizma
. Komparativna
fiziolo?ka
istra?ivanja diferencijacije “rasa” naj?e??e su obuhvatala karakteristi?ne elemente adaptacije na klimatske prilike, a posebno na temperaturu i
vla?nost
. Naga
osoba
(u mirovanju) toplotnu ravnote?u sa svojom okolinom uspostavlja na 27?30
0
C, a van toga raspona se uklju?uju odgovaraju?i homeostatski fiziolo?ki procesi. Iznad 30
0
C po?inje znojenje, a ispod 27
0
C tjelesna temperatura se reguli?e
drhtanjem
. Ovakve fiziolo?ke posljedice ima i poja?ana fizi?ka aktivnost, a pokrivanjem i utopljavanjem se posti?u adekvatni kompenzacijski efekti. Eksperimentalno je dokazano da su
ameri?ki
crnci
, uz sli?nu
konstituciju
, istovjetne uvjete okoline i jednak fizi?ki rad, relativno izdr?ljiviji na sparini, a bijelci u vreloj i suhoj klimi. Na vrelom i vla?nom vremenu, crnci, uz ni?u tjelesnu temperaturu i
puls
, gube za oko 5% manje tjelesne mase po satu fizi?ke aktivnosti, dok je situacija obrnuta u vrelim i suhim pustinjskim uvjetima.
Pretjeran gubitak toplote na niskim
temperaturama
se spre?ava su?avanjem perifernih krvnih sudova, ?to smanjuje protok
krvi
u ko?i i ekstremitetima. Tako se hladi samo povr?ina tijela, a odr?ava normalna i stalna
temperatura
vitalnih unutra?njih organa. Na oko 23
0
C neodjeven
bijelac
, u prosjeku, ima temperaturu rektuma od oko 36
0
C, glave oko 34,5
0
C, trupa oko 34
0
C, ?aka oko 30
0
C i stopala oko 25
0
C. Kada se udovi rashlade do oko 5?10
0
C opticaj krvi se intenzivira, pa (u tolerantnim granicama spoljne temperature) ne dolazi do smrzavanja eksponiranih dijelova tijela. U tom pogledu, međutim, postoje izrazite međupopulacijske razlike. Zabilje?eno je, naprimjer, da neodjeveni
australski
domoroci mogu spavati i na temperaturama na kojima prosje?an bijelac dr??e i cvoko?e, iako ima primjetno toplije udove.
Vi?i bazni
metabolizam
Eskimima
omogu?uje produkciju vi?e toplote, a smatra se da je poja?an
krvotok
u uronjenim dijelovima tijela (u hladnu
vodu
) kod njih također
geneti?ki
determiniran. Sli?na pojava je zabilje?ena i kod pripadnika plemena
Alakaluf
(sa krajnjeg
juga
?ilea
), koji ?ive u surovim uvjetima hladne i vjetrovite
klime
, a koji bez vidljivih posljedica hranu skupljaju rone?i u vodi temperature oko 6?8
0
C.
Op?a
priroda
međusobnog odnosa i kvantifikacija uticaja
filogenetske
(
geneti?ke
) i
fiziolo?ke
(idio)adaptacije na karakteristi?ne komplekse
klimatskih
faktora, uop?eno gledaju?i, jo? uvijek su (zapanjuju?e) nedovoljno poznati.
Ostali oblici
adaptacije
su superficijelno te?e uo?ljivi, iako su mnogi od njih, nesumnjivo, imali zna?ajnu ulogu u kreiranju mnogih bitnih crta “rasne” i op?e međupopulacijske diferencijacije
ljudskih
grupa. Tako je neporeciva pretpostavka da je odabiranje putem razli?itih
bolesti
i njihovih
pandemija
moglo biti izuzetno efikasan kreator karakteristi?nog geneti?kog sastava određenih
populacija
. Relativno otporniji
genotipovi
su u adekvatnoj mjeri imali ve?e u?e??e u formiranju razli?itih
genskih
fondova, a zna?aj ovog faktora podvla?i i mogu?nosti da istovjetne geneti?ke osnove nisu bile favorizovane u razli?itim populacijama. Drugim rije?ima, isti geni na jednom kraju svijeta mogli su pove?avati adaptivnu vrijednost svojih nosilaca, dok su u drugom stvarali izrazite predispozicije za neku
bolest
. Tako, naprimjer, otpornost na
malarijsku
groznicu koindicira sa tamnom
pigmentacijom
, a razli?ita
ishrana
favorizira određene
tipove konstitucije
; selektivni ?inilac mo?e biti i nedostatak izvjesnih
mikroelemenata
u zemlji?tu (i ?ivotnim namirnicama) itd. Fizi?ke osobenosti
stanovni?tva
jugo
isto?ne
Azije
mogle bi biti u vezi sa dominacijom pirin?ane ishrane, a zdepasta i de?mekasta konstitucija, sklona
gojaznosti
, u
sjevernoj
Aziji
i nekim dijelovima
Evrope
je, vjerovatno, adaptabilnija na nepovoljnu i neravnomjernu dinamiku ishrane (koja podrazumijeva stvaranje
energetskih
rezervi). Mi?i?avost i ?ilavost također predstavljaju komparativnu prednost pripadnika
lova?kih
plemena
i sl. Pretpostavljena uloga
spolnog
odabiranja, prema veoma varljivom popularnom ukusu i ”ljepotnom idealu”, sasvim je neizvjesna, iako ga
seksualni
promiskuitet
(podr?avan ili prikriven) u svim oblicima socijalne organizacije, nesumnjivo, podr?ava.
Neadaptacijski faktori
, odnosno vanselekcijski
evolutivni
?inioci adaptivno vrijednijih
fenotipova
, također su mogli u zna?ajnoj mjeri u?estvovati u formiranju karakteristi?nih
genskih
fondova osobenih ljudskih skupina i
populacija
. Među njima je, svakako, potencijalno najefikasniji
geneti?ki drift
, koji (bez obzira na adaptivnu vrijednost) na nekoj od klju?nih etapa formiranja genskog fonda naredne generacije, sasvim slu?ajno, određuje relativno u?e??e pojedinih gena. Njegovim djelovanjem, posebno u uvjetima reproduktivne izolacije, obja?njavaju se ?esto drasti?ne međupopulacijske razlike u frekvenciji određenih ”adaptivno neutralnih
osobina
”, koje, obi?no, nastaju slu?ajnim formiranjem
genskog
fonda pojedinih
izolata
i
migriraju?ih
grupa. One po ”principu osniva?a” konstitui?u genske fondove novih (sub)
populacija
, geneti?kog sastava koji u manjoj ili ve?oj mjeri (ali gotovo redovno) odstupa od aktuelnog stanja u mati?noj populaciji. Između ostalog, tom ”igrom slu?aja” u zahva?enim malim populacijama mo?e do?i do fiksiranja izuzetno visoke ili niske frekvencije određenih gena, bez obzira na njihov po?etni doprinos adaptivnoj vrijednosti fenotipa. S tim u vezi potrebno se podsjetiti da isti
gen
mo?e biti koristan u kombinaciji sa jednim, neutralan sa drugim, a ?tetan u kombinaciji sa tre?im
genom
te da njihovi efekti nisu izlo?eni samo prilagođavanju spoljnoj sredini nego međusobnoj interakciji.
Prema tome, i
geneti?ki drift
i
izolacija
uti?u na formiranje karakteristi?nih skupina balansiranih sistema gena, koje, u daljem konstituiranju genskog fonda populacija, mogu biti izlo?eni veoma heterogenim kompleksima ?inilaca prirodnog odabiranja, a u pravcu pove?anja op?e adaptiranosti uvjetima ?ivotne sredine. Međutim, ve? je dokazano da temeljna i sredi?nja svojstva
ljudske
vrste
?
razum
i
inteligencija
? nisu ni “rasno” ni populacijski ograni?ena. Postoje, dakle, izvjesne populacije koje druge nadma?uju u prosje?noj tjelesnoj visini,
pigmentaciji
,
dlakavosti
ili u?estalosti
krvnih grupa
razli?itih sistema (npr.), ali nisu zabilje?ene takve koje bi bile superiorne po su?tinskim
ljudskim
potencijalima.
Kao i sve druge grupne ljudske kategorije (populacije,
plemena
,
klase
,
nacije
,
kaste
i druge zajednice), “rase” su dinami?ni sistemi u vremenu i
prostoru
? ?ire se i smanjuju, nastaju i nestaju, integri?u se i diferenciraju, mijenja se njihova
biolo?ka
i
kulturna
struktura. Otuda savremene “rasne”skupine u svom genskom i kulturnom fondu, u manjoj ili ve?oj mjeri, nose tragove međudjelovanja i pro?imanja sa svojim susjedima.
 "Rasni" diverzitet
azijskiog
stanovni?tva
(Nordisk familjebok,1904)
"Rasni" diverzitet
azijskiog
stanovni?tva
(Nordisk familjebok,1904)
Biasutti (
1955
.)
[17]
klasificira "rase" prema prihvatljivijim kriterijima, posebno respektiraju?i prostorno?
geografske
, mogu?e
filogenetske
i
geneti?ke
pokazatelje međugrupnih relacija. Na osnovu tih i nekih drugih polazi?ta, ovaj autor i njegovi sljedbenici diferenciraju velike primarne i sekundarne skupine “rasnih oblika”. Prema njima, velike primarne skupine su ekvatorska i borealna (sjeverna), a sekundarne ? subekvatorska i pacifi?no?ameri?ka. Unutar tih skupina oni razlikuju u?e kategorije: grana (4), loza (16), “rasa” (52) i “podrasa”. Ovdje ?e biti prikazani detalji njihovog originalnog opisa koji su, u najve?oj mjeri, asocijativni i u poređenjima sa ostalim klasifikacijama (koje slijede). Najprepoznatljivije op?e odlike “rasnih grana” (primarni oblici) i loza (sekundarni oblici) izlo?ene su tabelarno, kako slijedi.
"Rasna skupina" primarnih ekvatorskih oblika (negro-australoidna)
[
uredi
|
uredi kod
]
Ova velika "rasna skupina" obuhvata australoidnu i negroidnu granu, sa po tri loze i ukupno 19 “rasa”.
Pretpostavlja se da su sve okeanijske populacije u srodni?koj vezi sa australoidima, a sve afri?ke ? sa negroidima, pri ?emu su australoidi najstarija i najprimitivnija skupina. Na osnovu nekih zajedni?kih obilje?ja, također se vjeruje da primarni ekvatorski oblici poti?u od prastanovni?tva (u pleistocenu) potonulog Sundajskog kopna. Prema jednoj drugoj hipotezi, međutim, centar njihove adaptivne radijacije mogao je biti negdje u ju?noj
Indiji
.
| Prostorni i filogenetski odnosi recentnih ljudskih populacija
|
| "Rasne" skupine
|
Grane
|
Loze
|
''Rase''
|
Primarni ekvatorski oblici
(1-2)→
Primarni borealni oblici
(3-4)→
Sekundarni subekvatorski oblici
(Paleoindidi, Indidi)→
Sekundarni pacifi?ko?ameri?ki oblici
(Polinezidi, Amerikanidi)→
|
1.
Australoidi
(Australidi, Papuanidi, Vedidi)→
2.
Negroidi
(Steatopigidi, Pigmidi, Negridi)→
3.
Mongoloidi
(Paleomongolidi, Mongolidi, Eslimidi)→
4.
Evropeidi
(Paleevropidi, Evropidi)→
5.
Amerikanidi
→
|
Australidi
(1-3)→
Papuanidi
(4-6)→
Vedidi
(7-8)→
Steatopigidi / Koisanidi
(9-10)→
Pigmidi
(11-12)→
Negridi
(13-19)→
Palemongolidi
(20-23)→
Mongolidi
(24-26)→
Eskimidi
(27)→
Paleevropidi
(28-29)→
Evropidi
(30-38)→
Lapidi
(39)→
Paleoindidi
(40-41)→
Etiopidi
(42-44)→
Polinezidi
(45)→
Amerikanidi
(46-52)→
|
1. Australska
2. Tasmanijska
3. Neokaledonska
4. Papuo?melanezijska
5. Papuo?montana
6. Tapirska
7. Vedska
8. Malajska
9. Bu?mani
10 Hotentoti
11. Bambuti
12. Babinga
13. Sudanska
14. Niloti
15. Kafridi
16. Silvidi
17. Batua
18. Andamanska
19. Semang (Negritos)
20. Paleosibirska
21. Tibetska
22. Punan
23. Tunguska
24. Kineska (Sinidi)
25. Ju?nomongolska
26. Eskimi
27. Ainu
28. Uralska
29. Mediteranska
30. Alpska
31. Baltijska
32. Jadranska (Dinarska)
33. Nordijska
34. Iranska (Orijentalna)
35. Indijska
36. Pamirska (Turanidi)
37. Laponska
38. Tamilska
39. Malabarska
40. Etiopska
41. Saharska
42. Malga?ka
43. Polinezijska
44. Aleganska
45. Dakota
46. Aleutska
47. Sonorska (Margidi)
48. Pueblo?andska
49. Amazonska (Brazilidi)
50. Lagidi
51. Pampidi (Patagonci)
52. Mageljanska (Fuegidi)
|
Veliku “rasnu skupinu” primarnih sjevernih oblika ?ine dvije grane (mongoloidi i evropeidi) sa po tri loze i ukupno 18 “rasa”
Proces specijalnog i temporalnog diferenciranja, odnosno adaptivne radijacije, borealnih “rasnih oblika” nedovoljno je poznat. Pretpostavlja se da su op?e klimatske prilike i stupanj insolacije bili osnovni faktori diversifikacije njihovih specifi?nih obilje?ja (kao ?to su uske nosnice, osobenost konstitucija, pigmentacija i neka druga). Dosada?nji nalazi ne potvrđuju mogu?nost da su geografske barijere (
Himalaji
,
Tien?an
,
Pamir
,
Altaj
) u najve?oj mjeri uticale na razdvajanje primarno homogenog borealnog prastanovni?tva na isto?ni (mongoloidni) i zapadni (evropeidni) dio. Prema jednoj od prihvatljivih hipoteza, ove skupine su se tokom posljednje
glacijacije
izdiferencirale na azijski i evropski centar radijacije njihovih “rasnih oblika”.
- Mongoloidi
obuhvataju loze
paleomongolida
,
mongolida
i
eskimida
.
Paleomongolidi
uklju?uju
paleosibirsku
, odnosno njenu savremenu “podrasu”
kam?adala
(
Samojedi
,
?uk?i
, Keti i Oroki ili Nanajci),
tibetsku
(
Tibet
, jugozapadna
Kina
,
Burma
) i
punan
(
Borneo
,
Sijam
,
Malaja
) “rasu”.
Mongolidi
, uz op?e mongoloidne odlike, nagla?enije imaju ?u?kastu ko?u, glatku, crnu i tvrdu kosu te jako izra?ene kosti lica. Srednjeg su ili ni?eg rasta i mezocefalni, a mongolski nabor (iznad o?iju) je izrazit. Ovdje spadaju tri “rase”:
tunguska
(
Mand?urija
, isto?ni
Sibir
,
Mongolija
i
Turkestan
),
kineska
(
sinidi
:
srednjokineska
i
ju?nokineska
“podrasa”) i
ju?nomongolska
(ju?na
Kina
,
Burma
,
Sijam
,
Indokina
).
Eskimidi
i njihova jedina “rasa” ?
Eskimi
naseljavaju obale
ameri?kog
Arktika
.
- Evropeidi
imaju tri loze:
paleevropidi
,
evropidi
i
lapidi
, sa ukupno 11 “rasa”.
Paleevropidi
su bili ?iroko rasprostranjena praskupina
evroazijskog
kopna, ?iji su
fosilni
tragovi registrovani od
Kine
(D?oukoudien) do zapadne
Evrope
(Combe-Capell,
Oberkassel
). Sada su predstavljeni samo
Ainu
“rasom” (ju?ni
Sahalin
,
Hokaido
,
Kurilski otoci
) i
uralskom
(sjeveroisto?na
Rusija
,
Altaj
, zapadni
Sibir
). Ainu su najdlakaviji ljudi dana?njice, a ponekad imaju nagla?en i nabor
o?nih
kapaka
- epikantus (“mongolske” o?i).
Evropidi
, pored op?ih evropeidnih oznaka, posebno se odlikuju relativno kratkim rukama, ?irokom karlicom, dobro razvijenom muskulaturom, veoma reduciranim aparatom za ?vakanje, uskim i prominentnim nosom i dobro oblikovanom bradom. Obuhvataju slijede?e “rase”:
mediteranska
(“podrase”:
berberska
,
paleosardska
i
litoralna
),
nordijska
(“podrase” ?
danska
,
finska
i
irska
),
iranska
ili
orijentalna
(“podrase” ?
asiroidska
i
libijska
),
indijska
(sjeverna
Indija
),
alpska
(srednja
Francuska
, ju?na
Njema?ka
,
?vicarska
, sjeverna
Italija
,
Austrija
,
?e?ka
i
Mađarska
),
baltijska
(“podrase” ?
paleoslavenska
i
karpatska
;
Poljska
,
Rusija
,
Finska
i Pri
baltik
),
jadranska
ili
dinarska
(“podrase”:
padska
i
nori?ka
; zapadni
Balkan
i
Karpati
do
Alpa
) i
pamirska
(
turanidi
:
armenoidska
“podrasa” iz
Male Azije
,
Irana
i
Armenije
).
Lapidi
, tj. njihova “rasa”
Laponci
, naseljavaju sjevernu
Skandinaviju
i poluotok
Kolu
. Imaju osebujne oznake, a među njima i takve po kojima li?e na pripadnike
alpske
“rase”.
Prema odgovaraju?im svojstvima ko?e, populacije ovih oblika li?e na ekvatorsko stanovni?tvo, a po op?em habitusu, konstituciji i strukturi tijela ? na borealne evropeide. Ovdje se svrstavaju loze paleoindida i etiopida (ukupno pet “rasa”.
- Paleoindidi
imaju osnovna paleoevropidska obilje?ja, koja su kombinovana sa australoidnim oblikom nosa i melanizmom. ?ine ih
tamilska
(jugoisto?na
Indija
) i
malabarska
“rasa” (jugozapadna
Indija
).
- Etiopidi
imaju evropeidni habitus sa nizom negroidnih primjesa. Obuhvataju
etiopsku
(“podrase”:
oromonska
,
Masai
i
Wa Tutsi
;
Etiopija
),
saharsku
(gornja Tibesti) i
malga?ku
(
Madagaskar
) “rasu”.
Ova skupina obuhvata veoma heterogene sekundarne “rasne oblike”, koji se svrstavaju u dvije loze: polinezidi (jedna “rasa”) i amerikanidi (devet “rasa”).
- Polinezidi
, odnosno polinezijska “rasa”, predstavljeni su
mikronezijskom
(Karolinski, Marijanski, Mar?alovi i Gilbertovi otoci) i
maorskom
(
Maori
) “podrasom” (
Novi Zeland
). Nastali su mije?anjem paleoevropidnih i paleomongolidnih elemenata, ali se po nizu izrazitih specifi?nosti smatraju posebnim i autohtono oblikovanim “rasnim tipom”.
- Amerikanidi
uklju?uju slijede?e “rase”:
aleganska
(
Njufaundlend
i jugoisto?na
Kanada
),
dakota
(
SAD
,
Kanada
i
Meksiko
),
aleutska
(sjeverozapad
Sjeverne Amerike
),
sonorska
ili
margidi
(
Meksiko
i
Kalifornija
),
pueblo?andska
(
Florida
,
Teksas
,
Meksiko
, Srednja
Amerika
,
Kolumbija
i
Venecuela
),
amazon
ska ili
brazil
idi (
Brazil
),
lagidi
(ju?ni
Brazil
),
pampidi
ili
patagonci
(
Urugvaj
i
Argentina
),
mageljanska
ili
fuegidi
(jugozapadni
Brazil
, sjeverozapadni i jugozapadni
?ile
).
U amerikanidnoj skupini “sekundarnih rasa” najupadljivija su paleomongolidna obilje?ja, na ?emu se i temelji op?eprihva?ena teza da su
Ameriku
(preko nekada?nje
Beringije
), u nekoliko talasa, naselili mongolidi. Sli?nost somatske strukture savremenih amerikanida i stanovni?tva
Okeanije
indicira mogu?nost da su ovo oto?je prvi naselili najstariji
borealni evropeidi
.
Na osnovu ve?ine uo?ljivih
morfolo?ko
?
anatomskih
komponenti ukupnog
fenotipa
, Garn (
1961
.) je izdvojio devet glavnih “geografskih rasa”:
amerikanidska
,
polinezidska
,
mikronezijska
,
malezijsko?papuanska
,
australijska
,
azijska
,
indijska
,
evropska
i
afri?ka
. Ne?to prije toga, međutim, Coon, Garn i Birdsell (
1950
.) predlo?ili su klasifikaciju od 30, a zatim Garn (
1961
.) od 32 “rase”; integracijom tih sistema nastala je tipologija sa 34 “rase”.
| Kun?Garn?Berdselova klasifikacija “rasa” i njihovo rasprostranjenje
[15]
|
| "Rasa"
|
Od
|
Do
|
| 1.
Sjevero
zapadna
evropska
|
Skandinavija
,
Sjeverna
Njema?ka
,
Sjeverna
Francuska
|
Beneluks
,
Velika Britanija
,
Irska
|
| 2.
Sjevero
isto?na
evropska
|
Poljska
,
Rusija
|
Ve?i dio
Sibira
|
| 3.
Alpska
|
Srednja
Francuska
, Ju?na
Njema?ka
,
?vicarska
, Sjeverna
Italija
|
Obala
Crnog mora
|
| 4.
Mediteranska
|
Tanger
, Obje strane
Mediterana
|
Dardaneli
,
Arabija
,
Turska
,
Iran
,
Turkmenija
|
| 5.
Hinduska
|
Indija
|
| 6.
Tunguska
|
Turkmenistan
|
| 7.
Tibetska
|
Tibet
|
| 8.
Sjeverno
-
kineska
|
Sjeverna
Kina
|
Srednja
Kina
|
| 9.
Klasi?na
mongoloidna
|
Sibir
|
Mongolija
|
| 10.
Eskimska
|
Ameri?ki
Arktik
|
| 11.
Jugo
isto?na
azijska
|
Ju?na
Kina
|
Tajland
,
Burma
,
Malaja
,
Indonezija
|
| 12.
Ainu
|
Sahalin
|
Sjeverni
Japan
(domoroci)
|
| 13.
Laponska
|
Arkti?ka
Skandinavija
|
Finska
|
| 14.
Sjeverno
ameri?ka
indijanska
|
Starosjedioci
SAD
|
Starosjedioci
Kanade
|
| 15.
Srednjo
ameri?ka
indijanska
|
Jugo
zapad
SAD
, Srednja
Amerika
|
Bolivija
|
| 16.
Ju?no
ameri?ka
indijanska
|
Peru
,
Bolivija
|
?ile
|
| 17.
Fuegidska
|
Ju?ni
dio
Ju?ne Amerike
|
| 18.
Isto?no
afri?ka
|
Isto?na
Afrika
,
Etiopija
|
Sudan
|
| 19.
Sudanska
|
Sudan
(ve?i dio)
|
| 20.
?umska
crna?ka
|
Zapadna
Afrika
|
Kongo
(ve?i dio)
|
| 21.
Bantu
|
Ju?na Afrika
|
Isto?na
Afrika
|
| 22.
Hotentotska
i
bu?manska
|
Ju?na Afrika
(domoroci)
|
Isto?na
Afrika
|
| 23.
Afri?ka
pigmejska
|
Ekvatorska
Afrika
(
tropske ki?ne ?ume
)
|
| 24.
Dravidska
|
Ju?na
Indija
|
Cejlon
|
| 25.
Negritska
|
Filipini
|
Andamani
,
Malaja
,
Nova Gvineja
|
| 26.
Melanezijsko
?
papuanska
|
Nova Gvineja
|
Oto?je
Fid?i
|
| 27.
Merinska
|
Jugo
isto?na
Australija
|
| 28.
Karpentarijska
|
Sjeverna
Australija
|
Srednja
Australija
|
| 29.
Mikronezijska
|
Otoci
zapadnog
Pacifika
|
| 30.
Polinezijska
|
Otoci srednjeg i isto?nog
Pacifika
|
| 31.
Neohavajska
|
Havaji
|
| 32.
Ladinska
|
Sjeverna Amerika
(nastaju?e
stanovni?tvo
)
|
Ju?na Amerika
(nastaju?e
stanovni?tvo
)
|
| 33.
Sjevernoameri?ka
“obojena”
|
Sjeverna Amerika
|
| 34.
Ju?noafri?ka
|
Ju?na Afrika
|
Najzna?ajnija diferencijalna crta ove, kao i Biasutijeve klasifikacije, u odnosu na one koje su prethodile, je prihvatanje ?injenice da su “rase”, ustvari, mendelovske
populacije
, podlo?ne mijenjanju, a ne apstraktni i nepromjenljivi
tipovi
i ”komponente”. Tako se razvoj najmanje ?etiri (od 34) “rase” mo?e pratiti ve? od po?etka njihovog formiranja (tokom proteklih pola milenija): sjevernoameri?ka “obojena rasa” (33) nastala je mije?anjem “rasa” pod rednim brojem 1, 3, 4, 20, 21, a vjerovatno i nekih drugih; ju?noafri?ka “obojena” (34) je, ustvari, mje?avina “rasa” 1, 3, 21 i 22; ladinska (32) ? od 4, 15, 16, 20 i 21, a neohavajska (31) ? od 1, 4, 8, 9, 11 i 30.
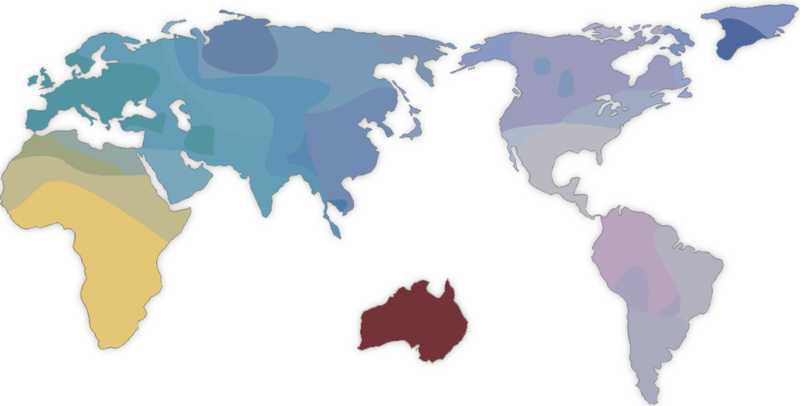 Karta svijeta na osnovu geneti?ke analize glavnih komponenti ljudskih populacija (iz knjige
The History and Geography of Human Genes
, 1994).
[18]
Karta svijeta na osnovu geneti?ke analize glavnih komponenti ljudskih populacija (iz knjige
The History and Geography of Human Genes
, 1994).
[18]
Schlaining-?ka deklaracija protiv rasizma, nasilja i diskriminacije
[
uredi
|
uredi kod
]
Međunarodna konferencija “Protiv rasizma, nasilja i diskriminacije” je odr?ana od 08. do 11. juna
1995
. godine u Stadtschlainingu, u organizaciji
Evropskog
univerzitetskog centra za mirovna istra?ivanja (EPU),
Austrijskog
UNESCO
komiteta, Instituta za humanu biologiju Univerziteta u
Be?u
, a u saradnji sa UNESCO?m i austrijskim Federalnim ministarstvom za
nauku
, istra?ivanja i
umjetnost
. U?esnici ove konferencije elaborirali su osnovne principe Schlaining?ke deklaracije. Na konferenciji su u?estvovali vode?i svjetski antropolozi i geneti?ari, uklju?uju?i i tada (i jo? uvijek) najkompetentnijeg Luiđija Luku Kavali-Sforcu (Luigi Luca Cavalli-Sforza,
1922
. -).
[19]
[20]
Deklarativni segment teksta prezentira rezultate dva glavna dijela Konferencije, koji su fokusirali probleme iz oblasti prirodnih nauka (II) i interdisciplinarne aspekte (III) glavne teme Konferencije. Operativni segment (IV) sadr?i strategije i akcije protiv rasizma, nasilja i diskriminacije. Schlaining?ka deklaracija je prilog Međunarodnoj godini tolerancije (
1995
.), koja je utvrđena na Generalnoj skup?tini UN-a, kada je UNESCO oficijelno i određen za vode?u organizaciju u ovoj problematici.
(1) UN?ovim instrumentima (Međunarodna konvencija i deklaracija UN-a o eliminaciji svih oblika rasne diskriminacije) te Deklaracijom UNESCO-a o rasama i rasnim predrasudama iz 1978. kao i u ?etiri prethodne deklaracije o rasnim pitanjima sve dr?ave i narodi su pozvani da se bore protiv rasizma i rasnih predrasuda te da podr?e eliminaciju svih formi rasizma svim raspolo?ivim sredstvima.
(2) Uprkos historijskom osporavanju, svjetskoj osudi i nau?nom dokazu zastarjelosti koncepta “rase”, “zlo rasizma” nije prevaziđeno. Naprotiv, po zavr?etku konflikta između Istoka i Zapada, rasizam je doveo do novih nasilja u vidu deportacija, masakra i poku?aja istrebljenja cijelih etni?kih grupa i manjina. Rasna diskriminacija, ekstremna desnica, ksenofobija, seksizam i netolerancija se ponovo ?ire. O?ivljavanje i eskalacija rasizma, njegovih nasilnih, novih i starih, pojavnih oblika, predstavlja jedan od glavnih izazova ?ovje?anstva. Za suo?avanje s tim bit ?e neophodno novo razumijevanje njegovih uzroka kao i razrada novih strategija i akcija.
(3) Objava iz Seville (1986), koja se odnosi na borbu protiv zloupotrebe rezultata biolo?kih istra?ivanja u legalizaciji rata i nasilja, odbacuje zabludu da je organizirano nasilje nad ljudima biolo?ki utemeljeno.
U svakom slu?aju, nau?no znanje nije dovoljno da odagna nasilje iz istorije. Broj ratova u siroma?nim i zemljama u razvoju od 1945. je u neprestanom porastu. U cjelini, broj ratova između dr?ava se smanjio, ali unutra?nji razdor i civilni ratovi prijete da postanu normalna pojava u perifernim i nerazvijenim regionima.
(4) Deklaracija iz Yamoussoukroa iz 1989. uzroke rastu?eg broja oru?anih sukoba nalazi u rasizmu, netoleranciji i diskriminaciji te poziva na podr?ku mirovnim programima i promociji mirovnih istra?ivanja. U isto vrijeme, za?eta je vizija kulture mira.
(5) UN rezolucija o godini tolerancije (1995.) zahtijeva stvaranje tolerantnog dru?tva i nagla?ava “…da tolerancija ? priznavanje i po?tivanje drugih, mogu?nost su?ivota i razumijevanja drugih ? predstavlja ?vrst temelj svakog civiliziranog dru?tva i mira”.
(6) Borba protiv rasne netolerancije i diskriminacije ne smije biti ograni?ena samo na moralnu osudu i javni prezir, ma koliko razumna i va?na bila ova reakcija kao prvi znak protesta. Saniranje simptoma i ubla?avanje posljedica rasizma, diskriminacije i nasilja, u smislu provođenja direktnih i brzih za?titnih mjera, kao i humanitarne pomo?i u hitnim slu?ajevima, je osnovni princip aktivne i zdru?ene humanitarnosti. U svakom slu?aju, kako bismo se uspje?no suo?ili i, jednom za svagda, pobijedili rasizam, nasilje i diskriminaciju, neophodno je sistematski na?i i eliminirati njihovo korijenje.
(7) Deklaracija iz Schlaininga, stoga, poziva na sveukupnu humanost, bez obzira na spol, zanimanje, socijalni status ili godine, da zajedno, solidarno djeluju protiv rasizma, nasilja i diskriminacije.
- II.
Zastarjelost koncepta "rase"
(8) Revolucija u na?em razumijevanju populacijske genetike i molekularne biologije je dovela do eksplozije saznanja o ?ivim bi?ima. Među idejama koje su su?tinski izmijenjene su i koncepti o ljudskoj varijaciji. Koncept “rase”, koji je iz pro?losti prenesen u dvadeseti vijek, potpuno je zastario. Uprkos toj ?injenici, koncept “rase” je kori?ten za opravdavanje povreda ljudskih prava. Va?an korak u prevenciji zloupotrebe geneti?kih argumenata je zamjena zastarjelog koncepta “rase” sa idejama i zaklju?cima baziranim na savremenom shvatanju geneti?ke varijacije primijenjenom na ljudskim populacijama.
(9) Tradicionalno se vjeruje da su “rase” geneti?ki homogene i da se međusobno razlikuju. Ova definicija je razvijena kako bi opisala ljudski diverzitet povezan s, na primjer, razli?itim geografskim lokalitetima. U svakom slu?aju, skora?nji napredak u modernoj biologiji, baziran na tehnikama molekularne genetike i matemati?kim modelima populacijske genetike, pokazao je da je ova definicija totalno neadekvatna. Aktuelna nau?na otkri?a ne podr?avaju raniju klasifikaciju ljudske populacije na posebne “rase” kao ?to su “Afrikanci”, “Evroazijati” (uklju?uju?i “Ameri?ke Indijance”) ili na ve?i broj subdivizija.
(10) U specifi?nim slu?ajevima mogu?e je utvrditi geneti?ke razlike između ljudskih populacija i manjih grupacija. Ove geneti?ke razlike pokazuju tendenciju pove?anja sa pove?anjem geografske udaljenosti, ali bazi?na geneti?ka varijacija između populacija je daleko manje istaknuta. To zna?i da je geneti?ki diverzitet ljudi postepen i ne pokazuje zna?ajan diskontinuitet između ljudskih populacija. Otkri?a koja potkrepljuju ovaj zaklju?ak prkose tradicionalnoj klasifikaciji “rasa” i ?ine svaki tipolo?ki pristup neprimjerenim.
Nadalje, molekularne analize razli?itih multialelnih gena pokazuju da je nasljedna varijacija znatna unutar bilo koje grupe, dok je, s druge strane, posmatrana međugrupna varijacija zanemarljivo mala.
(11) Razlike u vanjskom izgledu (boja ko?e, morfologija tijela i lica, pigmentacija itd.) među ljudima iz razli?itih dijelova svijeta je lahko zamjetljiva. Iako izgleda paradoksalno, ako uzmemo u obzir postojanje vidljivih, geneti?ki određenih morfolo?kih razlika, geneti?ka varijacija u fundamentalnim fiziolo?kim karakteristikama i funkcijama je zanemarljiva kada se uzmu u obzir populacioni prosjeci. Drugim rije?ima, percipiranje morfolo?kih razlika mo?e nas dovesti do pogre?nog zaklju?ka o su?tinskim geneti?kim razlikama.
(12) Dokazi pokazuju da je tokom evolucije savremenog ?ovjeka bilo relativno malo promjena u osnovnoj geneti?koj konstituciji populacija. Molekularne analize gena, također, sna?no sugeriraju da se savremeno ?ovje?anstvo tek nedavno pro?irilo u danas naseljena podru?ja svijeta, prilagođavaju?i se, tokom tog procesa, na veoma razli?ite i ponekad ekstremne uvjete sredine (npr. surove klime), i to u veoma kratkom vremenskom periodu. Neophodnost brzog prilagođavanja na ekstremne ekolo?ke razlike je dovela do promjena u maloj garnituri gena koji određuju osjetljivost na faktore ?ivotne sredine. Va?no je spomenuti da su ove adaptacije kao odgovor na uvjete sredine u velikoj mjeri historijske i nisu zna?ajne za ?ivot u modernoj civilizaciji. Ipak, one se ponekad tuma?e kao odraz su?tinskih razlika između grupa ljudi, doprinose?i time konceptu “rase”.
(13) Prema nau?nom poimanju, kategorizacija ljudi na osnovu distribucije geneti?ki determinisanih faktora je vje?ta?ka i ohrabruje stvaranje beskrajne liste proizvoljnih i obmanjivih dru?tvenih vizija i poimanja. Osim toga, ne postoji uvjerljiv dokaz za "rasnu" divergenciju u intelektualnim, emocionalnim, motivacijskim ili bilo kojim drugim psiholo?kim ili bihevioralnim karakteristikama koje su nezavisne o kulturalnim faktorima. Dobro je poznato da određene geneti?ke osobine korisne u jednoj ?ivotnoj situaciji mogu biti nedostatak u drugoj.
(14) Rasizam je ubjeđenje da se ljudske populacije razlikuju po nasljednim osobinama dru?tvenog zna?aja time ?ine?i određene grupe superiornim ili inferiornim u odnosu na druge. Ne postoji uvjerljiv nau?ni dokaz da je ovo uvjerenje valjano. Ovim dokumentom se tvrdi da ne postoji nau?no utemeljen na?in karakterizacije ljudske raznolikosti upotrebom krutih termina “rasnih” kategorija ili tradicionalnog koncepta “rase”. Dakle, ne postoji nau?no opravdan razlog za dalju upotrebu termina “rasa”.
- ↑
1,0
1,1
Cavali-Sforza L. L., Bodmer W. F. (1999): The genetics of human populations. Dover PublicTIONS, Inc., Mineola, New York,
ISBN
0-486-40693-8
.
- ↑
2,0
2,1
Cavalli-Sforza L. L., Menozzi P., Piazza A. (1994): The history and geography of human genes. Princeton University Press, Princeton,
ISBN
0-691-02905-9
.
- ↑
3,0
3,1
Hiernaux J. (1969): Egalite ou inegalite des races? Editions Hachette, Paris.
- ↑
https://link.springer.com/chapter/10.1007/978-1-4684-9063-3_14
- ↑
https://www.jstor.org/stable/27503827
- ↑
Adler A. (1957): Understanding Human Nature. Premier Books, Fawcett.
- ↑
Biasutti R. (1959 ): Le razze e i popoli della terra. 1. Razze, popoli e culture. 2. Europa, Asia. 3. Africa. 4. Oceania, America, indice generale (3. riv. e aggiornata). UTET, Torino.
- ↑
Boyd W. C. (1950): Genetics and the Races of Man ? An Introduction to Modern Physical Anthropology. Little, Brown & Co., Boston.
- ↑
Coon C. S., Garn S. M., Birdsell J. B. (1950): Races. Thomas, Springfield.
- ↑
Mayr E. (1988): Toward a New Philosophy of Biology ? Observations of an Evolutionist. Harvard University Press, Cambridge, Mass.
- ↑
Back L., Solomos J., Eds (2002): Theories of race and racism: A Reader. Taylor & Francis, New York,
ISBN
020300597X
;
ISBN
9780203005972
.
- ↑
Cavalli-Sforza L. L., Menozzi P., Piazza A. (1994): The history and geography of human genes. Princeton University Press, Princeton,
ISBN
0-691-02905-9
.
- ↑
Adams M. et al (2013): Readings for diversity and social justice, 3rd Edition. Routhledge, Taylor & Francis Group, New York,
ISBN
978-0415892940
,
ISBN
0415892945
.
- ↑
Had?iselimovi? R. (2005): Bioantropologija ? Biodiverzitet recentnog ?ovjeka. Institut za geneti?ko in?enjerstvo i biotehnologiju (INGEB), Sarajevo,
ISBN
9958-9344-2-6
.
- ↑
15,0
15,1
Dobzhansky T. (1962): Mankind Evolving ? The Evolution of the Human Species. Yale University Press, New Haven and London.
- ↑
Mayr E. (1970): Populations, Species, and Evolution. Belknap Press of Harvard University Press, Cambridge, Mass.
- ↑
Biasutti R. (1959 ): Le razze e i popoli della terra. 1. Razze, popoli e culture. 2. Europa, Asia. 3. Africa. 4. Oceania, America, indice generale (3. riv. e aggiornata). UTET, Torino.
- ↑
Cavalli-Sforza, Luigi Luca (1994).
The History and Geography of Human Genes
. Princeton University Press. str.
136
.
ISBN
0691087504
.
- ↑
en.wikipedia.org/wiki/Luigi_Luca_Cavalli-Sforza
- ↑
it.wikipedia.org/wiki/Luigi_Luca_Cavalli-Sforza
- Race: the Power of an Illusion
companion website to California Newsreel feature, 2003, PBS
- James, Michael (2008)
"Race"
,
Stanford Encyclopedia of Philosophy
.
- "Understanding Race"
, American Anthropological Association's educational website, with links for primary school educators and researchers
- "Human races"
,
Encyclopædia Britannica Online.
- "The Myth of Race"
Arhivirano
2009-01-01 na
Wayback Machine-u
,
Medicine Magazine,
2007
.
- Is Race "Real"?
, forum by Social Science Research Council, includes
A.M. Leroi, 2005 op-ed article,
New York Times
Arhivirano
2013-03-31 na
Wayback Machine-u
, advocating biological conceptions of race; responses from scholars in various fields
More from Leori with responses
- Richard Dawkins (2004):
"Race and creation"
Arhivirano
2005-02-21 na
Wayback Machine-u
, (extract from
The Ancestor's Tale: A Pilgrimage to the Dawn of Life
), in
Prospect Magazine
.