Diplodocus
(
gr.
"doble viga"), tambien castellanizado
diplodoco
, es un
genero
con tres especies conocidas de
dinosaurios
sauropodos
diplodocidos
, que vivieron a finales del
periodo
Jurasico
, hace aproximadamente entre 155,7 y 145 millones de anos, en el
Kimmeridgiense
y el
Titoniense
, en lo que hoy es
Norteamerica
.
[
1
]
Los primeros
fosiles
fueron descubiertos en 1877 por
S. W. Williston
.
Vivio en lo que es hoy el oeste de Norteamerica durante el periodo
Jurasico Superior
. El
Diplodocus
es uno de los dinosaurios mas comunes de la parte superior de la
Formacion Morrison
, una secuencia de sedimentos marinos y aluvionales depositados hace aproximadamente 152 millones de anos. La Formacion Morrison muestra un ambiente dominado por gigantescos sauropodos como
Camarasaurus
,
Barosaurus
,
Apatosaurus
y
Brachiosaurus
, ademas del
Diplodocus
.
[
2
]
El
Diplodocus
esta entre los dinosaurios mas facilmente identificables, con su forma clasica, cuello y cola largos, y cuatro patas robustas. Durante muchos anos, era el dinosaurio mas grande conocido. Su gran tamano pudo haber sido un impedimento para los depredadores
Allosaurus
y
Ceratosaurus
, cuyos restos se han encontrado en los mismos
estratos
, lo que sugiere que coexistieron con el
Diplodocus
.
Etimologia
[
editar
]
El nombre generico
Diplodocus
es la forma en
neolatin
derivada del griego
διπλ?ο?
(
diploos
) ≪doble≫ y
δοκ??
(
dokos
) ≪viga≫, en referencia a la forma de los
huesos en chevron
a lo largo de la parte inferior de la cola. El termino fue cunado por Othniel Charles Marsh en 1878, pensando que esta forma de los huesos en chevrone era exclusiva de los Diplodocus.
[
3
]
[
4
]
Posteriormente, sin embargo, fue descubierta en otros miembros de la familia de los diplodocidos y en sauropodos no diplodocidos como Mamenchisaurus.
[
cita requerida
]
Descripcion
[
editar
]
 Tamano de
Diplodocus carnegii
(verde) y
D. hallorum
(naranja), comparados con un humano.
Tamano de
Diplodocus carnegii
(verde) y
D. hallorum
(naranja), comparados con un humano.
Los
Diplodocus
estan entre los dinosaurios mejor conocidos. Fueron
animales
cuadrupedos
muy grandes, de cuello largo y con una extensa cola en forma de
latigo
. Sus miembros delanteros eran ligeramente mas cortos que los traseros, lo que configura una postura predominantemente horizontal. La
mecanica
de estos animales de cola y cuello largos, con cuatro patas macizas, ha sido comparada con la de un
puente colgante
.
[
5
]
El especimen tipo de
Diplodocus carnegii
,
CM 84
es el mas completo hallado de su genero, estimado en alrededor de 25 metros de largo
[
6
]
y 6.5 m de alto al cuello.
[
7
]
[
5
]
[
8
]
Para la especie de
D. carnegii
, conocido por restos muy bien preservados. las estimaciones de su
masa
se situan en el rango de 11,5 a 19,7
toneladas
con los siguientes valores segun los autores 11.5 toneladas,
[
9
]
12.7 toneladas,
[
10
]
16 toneladas,
[
11
]
y 19,7 toneladas.
[
12
]
Las estimaciones de masa modernas para
Diplodocus carnegii
tienden a estar en el rango de 10 a 16 toneladas metricas.
[
8
]
[
10
]
[
13
]
[
14
]
[
15
]
Aunque dinosaurios como el
Supersaurus
eran probablemente mas largos, los restos fosiles hallados de estas otras especies son solo parciales.
[
16
]
Los restos parciales de
D. hallorum
han aumentado la longitud estimada del genero, aunque no tanto como se penso previamente. Cuando fue descrito en 1991, el descubridor, David Gillete, calculo que pudo haber medido hasta 52 metros de largo,
[
17
]
haciendole el dinosaurio mas largo conocido, excepto los dudosos dinosaurios pobremente conocidos como
Amphicoelias
. Algunas estimaciones del peso se extendieron tanto como hasta 113 toneladas. La longitud estimada inicial fue posteriormente revisada a la baja a 33,5 metros y luego a 32 metros.
[
18
]
[
19
]
Una revision mas reciente demuestra que las vertebras gigantes de la cola estaban colocadas realmente mas adelante en la cola que lo que D. Gillete las habia ubicado originalmente. Gillette habia colocado originalmente las vertebras 12-19 como vertebras 20-27. El estudio demuestra que el esqueleto completo del
Diplodocus
en el
Museo Carnegie de Historia Natural
de
Pittsburgh
,
Pensilvania
, en el cual las estimaciones de
Diplodocus (Seismosaurus) hallorum
se basaron, incluia la decimotercera vertebra de la cola perteneciente a otro animal, lo cual redujo las estimaciones del tamano de
D. hallorum
en alrededor del 30 %, quedando en aproximadamente 32 metros de largo y 25-30 toneladas.
[
6
]
Cuello y cabeza
[
editar
]
 a) craneo, b) clasica representacion de la cabeza con las fosas nasales en lo alto de la misma, c) teoria de Bakker sobre una trompa, d) moderna representacion con las fosas nasales abajo en el hocico y una posible camara de resonancia.
a) craneo, b) clasica representacion de la cabeza con las fosas nasales en lo alto de la misma, c) teoria de Bakker sobre una trompa, d) moderna representacion con las fosas nasales abajo en el hocico y una posible camara de resonancia.
Su cuello estaba formado por al menos 15
vertebras
, por lo que se cree que mantenian el cuello paralelo al suelo y que eran incapaces de elevarlo significativamente, no mas de 30 grados por encima de su eje horizontal.
[
20
]
Nunca se ha encontrado un craneo del que se pueda decir con certeza que pertenece al
Diplodocus
, aunque son bien conocidos los craneos de otros diplodocidos estrechamente relacionados con
Diplodocus
, como
Galeamopus
. Los craneos de los diplodocidos eran muy pequenos en comparacion con el tamano de estos animales.
Diplodocus
tenia pequenos dientes en forma de "clavija" que apuntaban hacia adelante y solo estaban presentes en las secciones anteriores de las mandibulas.
[
21
]
Su caja craneana era pequena como en otros diplodocidos.
La cabeza de los
Diplodocus
ha sido ampliamente representada con las fosas nasales en la parte superior de la misma, debido a la posicion de las aberturas nasales en el apice del craneo. Ha habido especulaciones acerca de si tal configuracion significa que los
Diplodocus
tuvieron una trompa.
[
22
]
Pero un estudio reciente
[
23
]
afirmo que no hay evidencias paleoneuroanatomicas que respalden la hipotesis de la trompa. Se ha senalado que el
nervio facial
de animales como el
elefante
es largo, ya que inerva una trompa. Las pruebas sugieren que el nervio facial es muy pequeno en los
Diplodocus
. Estudios de Lawrence Witmer publicados en 2001 indicaron que, mientras que las aberturas nasales estaban arriba en la cabeza, las actuales fosas nasales carnosas estaban situadas mucho mas abajo, en el hocico.
[
24
]
 Diplodocus carnegii
, parte de la coleccion del
Museo de La Plata
.
Diplodocus carnegii
, parte de la coleccion del
Museo de La Plata
.
Los
Diplodocus
tenian una cola extremadamente larga, compuesta de mas de 80 vertebras
caudales
,
[
25
]
cantidad que practicamente duplica el numero con el que contaban algunos sauropodos primitivos, como el
Shunosaurus
con 43, y que tambien supera a la que tenian
macronarianos
contemporaneos, como el
Camarasaurus
con 53. Existen especulaciones acerca de que los diplodocos podrian haber utilizado su cola como defensa
[
26
]
o para hacer ruido como un
latigo
.
[
27
]
Tambien podria haber servido como contrapeso para el cuello. La parte media de la cola tenia "dobles vigas",
huesos
cheurones
de forma extrana que le dan a los
Diplodocus
su nombre. Estas pudieron haber servido para proporcionar apoyo para las vertebras, o quizas para evitar que los
vasos sanguineos
se aplastasen en caso de que la pesada cola del animal chocase contra la tierra o contra algun temerario depredador. Estas "dobles vigas" tambien aparecen en algunos dinosaurios emparentados con los
Diplodocus
.
[
28
]
Miembros
[
editar
]
Como otros sauropodos, la mano o "pie delantero" del
Diplodocus
estaba altamente modificada, con los huesos del dedo y de mano dispuestos en una columna vertical, con una seccion de corte con forma de
herradura
. El
Diplodocus
carecia de garras a excepcion del dedo 1 de la mano, que eran inusualmente grandes comparada a la de otros sauropodos, aplanada de lado a lado, y separados de los huesos de la mano. La funcion de esta garra inusualmente especializada es desconocida.
[
29
]
Otros aspectos anatomicos
[
editar
]
Recientes descubrimientos han demostrado que los
Diplodocus
y otros diplodocidos pudieron haber tenido unas estrechas y puntiagudas espinas de
queratina
alrededor del
dorso
, como las de una
iguana
, de hasta 18
centimetros
de alto en su cola y posiblemente a lo largo de la espalda y el cuello tambien, como en los
hadrosauridos
.
[
30
]
[
31
]
Esta caracteristica, radicalmente diferente de la imagen que se tenia de estos animales, ha sido incorporada en recientes reconstrucciones, como en la miniserie
Walking with Dinosaurs
.
[
32
]
La descripcion original de las espinas informo que los especimenes en la Cantera Howe cerca de Shell, Wyoming estaban asociados con restos oseos de un diplodocido no descrito "parecido a
Diplodocus
y
Barosaurus
.
[
30
]
Desde entonces, los especimenes de esta cantera han sido referidos a
Kaatedocus siberi
y Barosaurus sp., En lugar de
Diplodocus
.
[
33
]
[
34
]
Descubrimiento e investigacion
[
editar
]
 Diplodocus
en el
Museo Nacional de Historia Natural del Instituto Smithsoniano
.
Diplodocus
en el
Museo Nacional de Historia Natural del Instituto Smithsoniano
.
Varias especies de
Diplodocus
fueron descritas entre 1878 y 1924. El primer esqueleto fue hallado en el ano 1878 en
Como Bluff
,
Wyoming
, por
Benjamin Mudge
y
Samuel Wendell Williston
. Fue el
paleontologo
Othniel Charles Marsh
quien le puso el nombre de
Diplodocus longus
, "doble viga larga", ese mismo ano.
[
35
]
Desde entonces, restos de
Diplodocus
han sido encontrados en la Formacion de Morrison, al oeste de
Estados Unidos
, en
Colorado
, en
Utah
,
Montana
y
Wyoming
. Los fosiles de estos dinosaurios son relativamente comunes, a excepcion del
craneo
que nunca se ha encontrado completo.
Los dos generos de sauropodos de la Formacion de Morrison,
Diplodocus
y
Barosaurus
, comparten huesos muy similares en sus miembros. En el pasado, muchos huesos aislados fueron automaticamente atribuidos a los
Diplodocus
pero pudieron, de hecho, haber pertenecido al
Barosaurus
.
[
36
]
Diplodocus longus
, la
especie tipo
original , se conoce a partir de dos vertebras caudales fragmentarias completas y varias mas de la Formacion Morrison en la Cantera Felch de Colorado. Aunque varios especimenes mas completos se han atribuido a
D. longus
,
[
37
]
analisis detallados han sugerido que el fosil original carece de las caracteristicas necesarias para permitir la comparacion con otros especimenes. Por esta razon, se ha considerado un
nomen dubium
, que no es una situacion ideal para la especie tipo de un genero conocido como
Diplodocus
. Se esta considerando una peticion a la
Comision Internacional de Nomenclatura Zoologica
que propone hacer que
D. carnegii
la nueva especie tipo.
[
33
]
[
38
]
D. lacustris
es una especie
dudosa
, nombrada por Marsh en 1884, a partir de restos de un pequeno animal proveniente de
Morrison
, Colorado.
[
39
]
En la actualidad se cree que estos huesos pertenecieron a un animal inmaduro, en vez de una especie separada.
[
40
]
En 2015, se concluyo que el especimen en realidad pertenecia a
Camarasaurus
[
33
]
D. carnegii
, tambien escrito
D. carnegiei
, llamado asi por
Andrew Carnegie
, es el mas conocido, principalmente debido a un esqueleto casi completo, especimen
CM 84
coleccionado por
Jacob Wortman
, del
Museo Carnegie de Historia Natural
en
Pittsburgh
,
Pensilvania
y fue descrito y nombrado por
John Bell Hatcher
en 1901.
[
41
]
[
42
]
Este fue considerado como especie tipo para
Diplodocus
por Tschopp en 2016.
[
38
]
Aunque no es la especie del holotipo, la mas conocida es
Diplodocus carnegie
, siendo la mas famosa debido al gran numero de exhibiciones que se han realizado de su esqueleto en museos de todo el mundo.
Diplodocus hayi
, conocido por un esqueleto parcial y craneo descubierto por
William H. Utterback
en 1902 cerca de
Sheridan
, Wyoming, fue descrito en 1924.
[
43
]
En 2015, fue renombrado como el genero separado
Galeamopus
y varios otros especimenes Diplodocus fueron referido a ese genero, sin dejar conocidos calaveras de
Diplodocus
.
[
33
]
Diplodocus hallorum
fue descrito por primera vez en 1991 por Gillette como
Seismosaurus halli
de un esqueleto parcial que comprende vertebras, pelvis y costillas, especimen
NMMNH P-3690
, fue encontrado en 1979.
[
44
]
Como el nombre especifico honra a dos personas, Jim y Ruth Hall, George Olshevsky mas tarde sugirio enmendar el nombre como
S. hallorum
, usando el genitivo plural obligatorio, Gillette luego modifico el nombre,
[
17
]
cuyo uso fue seguido por otros, incluido Carpenter en 2006.
[
18
]
En 2004, una presentacion en la conferencia anual de la Sociedad Geologica de America argumento para que
Seismosaurus
sea considerado un
sinonimo mas moderno
de
Diplodocus
.
[
45
]
Esto fue seguido por una publicacion mucho mas detallada en 2006, que no solo renombro a la especie
Diplodocus hallorum
, sino que tambien apunto que podria ser la misma que
D. longus
.
[
46
]
La posicion de que
D. hallorum
debe considerarse como un especimen de
D. longus
tambien fue tomada por los autores de una redescripcion de
Supersaurus
, refutando una hipotesis previa de que
Seismosaurus
y
Supersaurus
eran lo mismo.
[
47
]
Un analisis de 2015 de las relaciones diplodocidos senalo que estas opiniones se basan en los especimenes mas completos referidos de
D. longus
. Los autores de este analisis concluyeron que esos especimenes eran de hecho la misma especie que
D. hallorum
, pero que el propio
D. longus
era un nomen dubium.
[
33
]
Clasificacion
[
editar
]
 Segun los estudios realizados, los
Diplodocus
se levantaban sobre sus dos patas traseras y estiraban su largo cuello para llegar a las hojas mas verdes ubicadas en la copa de los arboles.
Segun los estudios realizados, los
Diplodocus
se levantaban sobre sus dos patas traseras y estiraban su largo cuello para llegar a las hojas mas verdes ubicadas en la copa de los arboles.
Diplodocus
es el
genero tipo
y da su nombre a la familia
Diplodocidae
, a la cual pertenece.
[
39
]
Los miembros de esta familia, aunque de gran tamano, poseen una estructura mas delgada en comparacion con la de otros sauropodos, como los
titanosaurianos
y los
braquiosauridos
. Todos estan caracterizados por largos cuellos y colas y una postura horizontal, con los miembros delanteros mas cortos que los traseros. Los diplodocidos dominaron a finales del Jurasico en Norteamerica y posiblemente Africa
[
25
]
y parecen haber sido reemplazados por los
titanosauridos
durante el
Cretacico
.
Una subfamilia,
Diplodocinae
, fue creada para incluir a los
Diplodocus
y a sus parientes mas cercanos, incluyendo a
Barosaurus
.
Apatosaurus
, aunque contemporaneo, tiene un parentesco mas lejano, pero sigue siendo considerado un diplodocido aunque no un diplodocino, ya que es un miembro de la subfamilia
Apatosaurinae
.
[
48
]
[
49
]
Los generos
Dinheirosaurus
y
Tornieria
han sido tambien identificados como parientes cercanos de
Diplodocus
por algunos autores.
[
50
]
[
51
]
El grupo Diplodocoidea abarca a los diplodocidos, asi como los
dicreosauridos
,
rebaquisauridos
,
Suuwassea
,
[
48
]
[
49
]
Amphicoelias
[
51
]
y posiblemente
Haplocanthosaurus
.
[
52
]
Anteriormente se incluia a los
Nemegtosauridae
, hoy incluido dentro de
Lithostrotia
dentro de
Titanosauria
.
[
53
]
Este
clado
es el grupo hermano de los camarasauridos, braquiosauridos y titanosaurianos; los
Macronaria
.
[
54
]
[
52
]
El conjunto de estos taxones conforman
Neosauropoda
, el mas grande, diverso y exitoso grupo de dinosaurios
sauropodomorfos
.
Filogenia
[
editar
]
El siguiente
cladograma
se basa en el
analisis filogenetico
realizado por Whitlock en 2011, que muestra las relaciones de
Diplodocus
entre los otros generos asignados a la familia Diplodocidae:
[
55
]
Cladograma
de Diplodocidae realizado por Tschopp, Mateus y Benson en 2015, el que se realiza por especies, notese la ausencia de
D. longus
ya que
D. carnegii
es considerado el tipo del genero.
[
33
]
Paleobiologia
[
editar
]
Debido a la gran cantidad de restos esqueleticos,
Diplodocus
es uno de los dinosaurios mejor estudiados. Muchos aspectos de su estilo de vida han sido objeto de diversas teorias a lo largo de los anos.
[
28
]
Las comparaciones entre los
anillos esclerales
de diplodocinos y las aves y reptiles modernos sugieren que pueden haber sido catemeral, activos a lo largo del dia a intervalos cortos.
[
56
]
Marsh y luego Hatcher supusieron que el animal era acuatico, debido a la posicion de sus aberturas nasales en el apice del craneo.
[
57
]
Comportamiento acuatico similar se describio comunmente para otros sauropodos grandes, como
Brachiosaurus
y
Apatosaurus
. Un estudio de 1951 de Kenneth A. Kermack indica que los sauropodos probablemente no podrian haber respirado por la nariz cuando el resto del cuerpo estaba sumergido, ya que la
presion
del agua en la pared del
torax
seria demasiado grande.
[
58
]
Desde la decada de 1970, el consenso general tiene los sauropodos como animales firmemente terrestres, navegando entre arboles, helechos y arbustos.
[
59
]
Los cientificos han debatido sobre como los sauropodos podian respirar con sus grandes cuerpos y largos cuellos, lo que habria incrementado la cantidad de espacio muerto . Probablemente tenian un sistema respiratorio aviar, que es mas eficiente que un sistema de mamiferos y reptiles. Las reconstrucciones del cuello y el torax de
Diplodocus
muestran una gran neumaticidad , que podria haber jugado un papel en la respiracion como lo hace en las
aves
.
[
60
]
 Antes, algunos cientificos creian que los
Diplodocus
caminaban con las extremidades flexionadas, pero en la actualidad, esta teoria ya ha sido descartada.
Antes, algunos cientificos creian que los
Diplodocus
caminaban con las extremidades flexionadas, pero en la actualidad, esta teoria ya ha sido descartada.
La representacion de la postura de los
Diplodocus
ha cambiado considerablemente con los anos. Por ejemplo, una obra clasica de 1910 de Oliver P. Hay representa a dos
Diplodocus
merodeando en la orilla de un
rio
. Dichos animales cuentan con miembros abiertos hacia los lados, similares a los de los lagartos. Argumento que los
Diplodocus
tenian un paso parecido al de un
lagarto
con patas sobresaliendo a ambos lados,
[
61
]
afirmacion que fue apoyada por Gustav Tornier. Sin embargo, esta hipotesis fue rebatida por William J. Holland, quien demostro que un
Diplodocus
con esa postura, habria necesitado un foso para hacer pasar su vientre.
[
62
]
 Representacion de un
Diplodocus
hecha por Oliver P. Hay en 1910.
[
63
]
Representacion de un
Diplodocus
hecha por Oliver P. Hay en 1910.
[
63
]
Mas adelante, los
diplodocidos
fueron retratados con frecuencia con sus cuellos elevados, lo que les permitiria comer de arboles altos, pero, recientemente, los cientificos argumentaron que el
corazon
habria tenido problemas para mantener la
presion arterial
necesaria para
oxigenar
el
cerebro
. Ademas, estudios posteriores comprobaron que la estructura de las vertebras
cervicales
no pudieron permitir que el cuello se moviese tan arriba. Los estudios que analizan la morfologia de los cuellos de sauropodo han concluido que la postura neutral del cuello
Diplodocus
era casi horizontal, en lugar de vertical, y cientificos como Kent Stevens lo han usado para argumentar que los sauropodos, incluido
Diplodocus
, no levantaron la cabeza demasiado por encima del hombro .
[
64
]
[
65
]
Un estudio de 2009 encontro que todos los
tetrapodos
parecen sostener la base de sus cuellos en la maxima extension vertical posible cuando se encuentra en una postura normal y alerta y argumento que lo mismo seria valido para los sauropodos, salvo caracteristicas desconocidas y unicas que distinguen la anatomia de los tejidos blandos de sus cuellos de otros animales. El estudio encontro fallas con las suposiciones de Stevens con respecto al rango de movimiento potencial en los cuellos de sauropodos, y con base en la comparacion de esqueletos con animales vivos, el estudio tambien argumento que los tejidos blandos podrian tener una mayor flexibilidad de lo que sugieren los huesos. Por estas razones, argumentaron que
Diplodocus
habria mantenido su cuello en un angulo mas elevado que los estudios previos habian concluido.
[
59
]
 Esqueleto de un
Diplodocus
con la postura reciente, expuesto en el
Museo de Historia Natural
de
Londres
.
Esqueleto de un
Diplodocus
con la postura reciente, expuesto en el
Museo de Historia Natural
de
Londres
.
Al igual que con el
Barosaurus
, el largo cuello de los
Diplodocus
es fuente de mucha controversia entre los cientificos. En 1992, un estudio de la
Universidad de Columbia
sobre la estructura del cuello de un diplodocido, indico que los cuellos tan largos habrian requerido un corazon de 1.6 toneladas. El estudio propuso que animales como este debieron tener "corazones" auxiliares rudimentarios en sus cuellos, cuyo unico proposito habria sido el de bombear la sangre al siguiente "corazon".
[
5
]
Alimentacion
[
editar
]
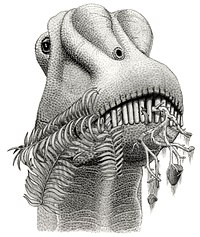 Un
diplodocido
comiendo helechos.
Un
diplodocido
comiendo helechos.
Comparados con los dientes de otros sauropodos, los del
Diplodocus
eran muy extranos. Las coronas dentales eran largas, delgadas y elipticas en seccion transversal, mientras que el apice forma un punto romo y triangular.
[
21
]
La faceta de uso mas prominente esta en el apice; sin embargo, a diferencia de los demas patrones de uso observados dentro de los sauropodos, los del
Diplodocus
se ubican en el lado
labial
, contra la
mejilla
, de los dientes superiores e inferiores.
[
21
]
Esto significa que los
Diplodocus
y otros diplodocidos tenian un mecanismo de alimentacion radicalmente diferente al de otros sauropodos. El deshojamiento de ramas usando un lado del hocico es la forma de alimentacion mas probable del
Diplodocus
,
[
66
]
[
67
]
[
68
]
ya que explica los poco habituales patrones de desgaste de los dientes a partir del contacto entre diente y comida. Al deshojar ramas usando un lado del hocico, una fila de dientes debio usarse para arrancar el follaje del tallo, mientras que la otra debio actuar como guia y estabilizador. Gracias a que contaba con una alongada region preorbital del craneo (delante de los ojos), podria deshojar porciones mas largas de tallos en una sola accion.
[
21
]
El movimiento hacia atras de las mandibulas inferiores pudo haber contribuido con dos significantes funciones en el comportamiento de alimentacion, aumentar la apertura de la boca y permitir ajustes finos en las posiciones relativas de las filas de dientes para afinar el deshoje.
[
21
]
La flexibilidad del cuello
Diplodocus
se debate, pero deberia haber sido capaz de navegar desde niveles bajos hasta aproximadamente 4 metros cuando esta en cuatro patas.
[
20
]
[
64
]
Sin embargo, los estudios han demostrado que el centro de masa de Diplodocus estaba muy cerca de la cavidad de la cadera.
[
69
]
[
70
]
esto significa que Diplodocus podria volver a una postura bipeda con relativamente poco esfuerzo. Tambien tenia la ventaja de utilizar su cola grande como un apoyo que permitiria una postura de tripode muy estable. En una postura de tripode,
Diplodocus
podria potencialmente aumentar su altura de alimentacion hasta aproximadamente 11 metros.
[
70
]
[
71
]
Con un cuello lateral y dorsoventralmente flexible y la posibilidad de utilizar su cola para erguirse apoyandose en sus miembros traseros, habilidad de ≪
tripode
≫, los
Diplodocus
habrian tenido la capacidad de alimentarse de hojas en muchos niveles, bajo, intermedio y alto, hasta aproximadamente 10 metros sobre el suelo, desde los pequenos
ginkgos
a las colosales
araucarias
.
[
72
]
El rango de movimiento del cuello tambien habria permitido que la cabeza pazca por debajo del nivel del cuerpo, lo que llevo a algunos cientificos a especular sobre si
Diplodocus
pastaba en las plantas acuaticas sumergidas, de las riberas de los rios. Este concepto de la postura de alimentacion se apoya en las longitudes relativas de las extremidades anteriores y posteriores. Ademas, sus dientes tipo clavija se pueden haber usado para comer
plantas acuaticas
blanda.
[
64
]
Matthew Cobley y colegas en 2013 disputan lo anteriormente dicho, encontrando que los musculos grandes y el cartilago darian movimientos limitados del cuello. Afirman que los rangos de alimentacion de los sauropodos como
Diplodocus
eran mas pequenos de lo que se creia anteriormente y los animales pueden haber tenido que mover sus cuerpos para acceder mejor a las areas donde pueden seleccionar la vegetacion. Como tal, podrian haber pasado mas tiempo alimentandose para satisfacer sus necesidades minimas de energia.
[
73
]
[
74
]
Las conclusiones de Cobley y colegas fueron disputadas en 2013 y 2014 por Mike Taylor, quien analizo la cantidad y posicion del cartilago intervertebral para determinar la flexibilidad del cuello de
Diplodocus
y
Apatosaurus
. Taylor descubrio que el cuello del
Diplodocus
era muy flexible y que Cobley y colegas estaban equivocados, en afirmar que la flexibilidad implicita en los huesos es menor.
[
75
]
Young y colegas en 2012 utilizaron modelos
biomecanicos
para examinar el rendimiento del craneo de
Diplodocus
. Se concluyo que la propuesta de que su denticion se usara para descortezar no fue respaldada por los datos, que mostraron que bajo ese escenario, el craneo y los dientes sufririan tensiones extremas. Se demostro que las hipotesis de mordida de ramas y mordeduras de precision son comportamientos de alimentacion biomecanicamente plausibles.
[
76
]
Los dientes tambien fueron reemplazados continuamente a lo largo de sus vidas, por lo general en menos de 35
dias
, como fue descubierto por Michael D'Emic y colegas. Dentro de cada alveolo dentario, se desarrollaron hasta cinco dientes de reemplazo para reemplazar el siguiente. Los estudios de los dientes tambien revelan que preferia una vegetacion diferente a la de los otros sauropodos del Morrison, como
Camarasaurus
. Esto puede haber permitido mejor que las diversas especies de sauropodos existan sin competencia.
[
77
]
En 2010, Whitlock y colegas describieron un craneo juvenil en el momento referido al
Diplodocus
,
CM 11255
, que diferia mucho de los craneos adultos del mismo genero, su hocico no era romo y los dientes no estaban confinados al frente del hocico. Estas diferencias sugieren que los adultos y los juveniles se alimentaban de manera diferente. Tal diferencia ecologica entre adultos y juveniles no se habia observado previamente en
sauropodomorfos
.
[
78
]
Al igual que otros
sauropodos
, a la hora de digerir los vegetales de los que se alimentaban se ayudaban de
gastrolitos
. La distribucion de los gastrolitos hallados junto a un fosil de
D. hallorum
, repartidos en dos conjuntos separados, podria sugerir que estos animales tenian un sistema digestivo similar al de las aves modernas, con un
buche
y una
molleja
, y que cada grupo de gastrolitos pertenecia a cada una de estas cavidades.
[
79
]
Reproduccion y crecimiento
[
editar
]
Si bien el cuello largo se ha interpretado tradicionalmente como una adaptacion de la alimentacion, tambien se sugirio que el cuello sobredimensionado de
Diplodocus
y sus parientes podria haber sido principalmente una exhibicion sexual, con cualquier otro beneficio de alimentacion en segundo lugar.
[
80
]
Un estudio de 2011 refuto esta idea en detalle.
[
81
]
Aunque no hay evidencia de habitos de anidamiento de los
Diplodocus
, otros
sauropodos
como el
titanosauriano
Saltasaurus
, han sido asociados con sitios de anidamiento.
[
82
]
[
83
]
Los sitios de anidamiento de los titanosaurianos, indican que pudo haber depositado sus huevos comunalmente a lo largo de una gran area en muchos hoyos bajos, cada uno de ellos cubierto con vegetacion. Esto ha generado la especulacion de que los
Diplodocus
pudieron presentar un comportamiento similar. Otra teoria es la inclusion de las hembras en zonas de
arboleda
mas frondosa, una vez fertilizadas, para la deposicion de los huevos en un lugar "mas seguro" que la desamparada planicie en la que habitualmente subsistian. El documental Walking with Dinosaurs retrato a una madre
Diplodocus
usando un
ovipositor
para poner huevos, pero fue pura especulacion por parte del autor del documental.
[
32
]
En
Diplodocus
y otros sauropodos, el tamano de las nidadas y los
huevos
individuales eran sorprendentemente pequenos para animales tan grandes. Esto parece haber sido una adaptacion a las presiones de predacion, ya que los huevos grandes requeririan un mayor tiempo de incubacion y, por lo tanto, estarian en mayor riesgo.
[
84
]
Segun varios estudios de
histologia
de
huesos
, los
Diplodocus
, al igual que otros sauropodos, crecian a un ritmo muy rapido, alcanzando la madurez sexual en tan solo una decada y continuando con su crecimiento a lo largo de toda su vida.
[
85
]
[
86
]
[
87
]
Esta postura es radicalmente distinta de la que se atribuia a los sauropodos, los cuales se pensaba que crecian lentamente durante su vida, tomando decadas para alcanzar su madurez.
Paleoecologia
[
editar
]
 Comparacion de dos modos de vida de
Diplodocus
propuestos a lo largo de la historia.
Comparacion de dos modos de vida de
Diplodocus
propuestos a lo largo de la historia.
La
Formacion Morrison
es una secuencia de sedimentos marinos y aluviales poco profundos que, segun la
datacion radiometrica
, oscila entre 156,3 millones de anos en su base,
[
88
]
y 146,8 millones de anos en la parte superior,
[
89
]
lo que lo situa en las etapas finales del
Oxfordiense
,
Kimmeridgiense
y
Titoniense
temprano del Periodo
Jurasico Superior
. Esta formacion se interpreta como un entorno
semiarido
con distintas estaciones humedas y secas. La cuenca de Morrison donde vivian los dinosaurios, se extendia desde
Nuevo Mexico
hasta
Alberta
y
Saskatchewan
y se formo cuando los precursores de la
Cordillera Frontal de las Montanas Rocosas
comenzo a empujar hacia el oeste. Los depositos de sus cuencas de drenaje orientadas al este fueron transportados por arroyos y rios y depositados en tierras bajas pantanosas, lagos, canales fluviales y llanuras aluviales.
[
90
]
Esta formacion es similar en edad a la
Formacion Lourinha
en Portugal y la
Formacion Tendaguru
en Tanzania .
[
91
]
La Formacion Morrison registra un ambiente y tiempo dominado por gigantescos dinosaurios sauropodos.
[
92
]
Los dinosaurios conocidos del Morrison incluyen los
teropodos
Ceratosaurus
,
Koparion
,
Stokesosaurus
,
Ornitholestes
,
Allosaurus
,
Saurophaganax
y
Torvosaurus
, los sauropodos
Apatosaurus
,
Brontosaurus
,
Brachiosaurus
,
Camarasaurus
y
Diplodocus
, y los ornitisquios
Camptosaurus
,
Dryosaurus
,
Othnielia
,
Gargoyleosaurus
y
Stegosaurus
entre otros.
[
93
]
Diplodocus
se encuentra comunmente en los mismos sitios que
Apatosaurus
,
Allosaurus
,
Camarasaurus
y
Stegosaurus
.
[
13
]
Allosaurus
representaba del 70 al 75% de los especimenes de teropodos y se encontraba en el nivel trofico superior de la red trofica de Morrison.
[
94
]
Muchos de los dinosaurios de la Formacion Morrison son del mismo genero que los que se ven en las rocas portuguesas de la Formacion Lourinha, principalmente
Allosaurus
,
Ceratosaurus
,
Torvosaurus
y
Stegosaurus
o tienen una contraparte cercana,
Brachiosaurus
y
Lusotitan
,
Camptosaurus
y
Draconyx
.
[
91
]
Otros vertebrados que compartio esta paleoambiente incluido
peces con aletas radiadas
,
ranas
,
salamandras
,
tortugas
como
Dorsetochelys
,
esfenodontos
,
lagartos
, terrestres y acuaticos
cocodrilomorfos
tales como
Hoplosuchus
y varias especies de
pterosaurios
como
Harpactognathus
y
Mesadactylus
y
mamiferos
primitivos
docodontes
,
multituberculados
,
symmetrodontes
, y
triconodontes
.. Conchas de
bivalvos
y
caracoles acuaticos
tambien son comunes. La flora del periodo ha sido revelada por fosiles de
algas verdes
,
hongos
,
musgos
,
colas de caballo
,
cicadas
,
ginkgoes
y varias familias de
coniferas
. La vegetacion variaba desde los bosques de helechos arborescentes y helechos,
bosques de galeria
, hasta las
sabanas
de helechos con arboles ocasionales, como la conifera
Brachyphyllum
de tipo Araucaria .
[
95
]
En la cultura popular
[
editar
]
 Presentacion de la primera replica de
Diplodocus carnegiei
a los miembros del
Museo de Historia Natural de Londres
, 12 de mayo de 1905. Lord Avebury aparece pronunciando un discurso.
Presentacion de la primera replica de
Diplodocus carnegiei
a los miembros del
Museo de Historia Natural de Londres
, 12 de mayo de 1905. Lord Avebury aparece pronunciando un discurso.
El
Diplodocus
es un dinosaurio famoso y muy representado dado que ha sido exhibido en mas lugares que ningun otro sauropodo. Mucho de esto se ha debido a la abundancia de restos esqueleticos y a haber sido considerado durante mucho tiempo como el dinosaurio mas largo. Replicas de los esqueletos de
Diplodocus
todavia se exhiben en muchos museos de todo el mundo, incluyendo un inusual
D. hayi
en el
Museo de Ciencias Naturales de Houston
. Al mismo tiempo, las donaciones realizadas por
Andrew Carnegie
a principios del siglo
XX
de numerosas copias en yeso de esqueletos completos de un
Diplodocus
, cuyo original es popularmente conocido como
Dippy
, desempenaron un papel de considerable importancia en la difusion internacional del conocimiento acerca de estos enigmaticos seres prehistoricos.
[
96
]
[
97
]
Tienen como minimo una de estas copias instituciones como el
Museo de Historia Natural de Londres
, el
Museo Nacional de Historia Natural de Francia
en
Paris
, el
Museo Nacional de Ciencias Naturales
en
Madrid
,
[
98
]
el
Museo de Historia Natural de Berlin
, el
Museo Senckenberg
(en
Francfort
, Alemania), el
Museo de Historia Natural
en la
Ciudad de Mexico
, el
Museo Field de Historia Natural
de
Chicago
, el
Museo de La Plata
en
Argentina
y, por supuesto, el original sigue siendo uno de los centros de atencion de las colecciones del
Museo Carnegie de Historia Natural
en Pittsburgh. Un esqueleto montado de
D. longus
se encuentra en el
Museo Nacional de Historia Natural de los Estados Unidos
en
Washington D. C.
, mientras que un esqueleto montado de
D. hallorum
(antes
Seismosaurus
), que puede ser el mismo que
D. longus
, puede ser encontrado en
Museo de Nuevo Mexico de Historia Natural y Ciencia
.
 Modelo de
Diplodocus
levantado sobre sus patas traseras en Bałtow, Polonia.
Modelo de
Diplodocus
levantado sobre sus patas traseras en Bałtow, Polonia.
Diplodocus
ha sido un tema frecuente en las peliculas del dinosaurio, documentales y ficticias. Fue mostrado en el segundo episodio de la serie de television premiada de la BBC;
Walking with Dinosaurs
. El episodio "La Era de los Titanes" sigue la vida de un
Diplodocus
hace 152 millones de anos, enfrentandose a ataques por parte de
Ornitholestes
,
Stegosaurus
y
Allosaurus
. La pelicula animada
Fantasia
ofrece muchos sauropodos en el segmento de ≪
La consagracion de la primavera
≫, uno de ellos parece ser un
Diplodocus
. En literatura,
James A. Michener
libro
Centennial
tiene un capitulo dedicado a
Diplodocus
, narrando la vida y la muerte de un individuo.
Diplodocus
es una figura que suele ser vista habitualmente entre los dinosaurios de
juguete
y entre las
maquetas
. Ha tenido dos modelos de juguete diferentes en la coleccion de juguetes Carnegie (la
Carnegie Collection
, vease aqui
el articulo correspondiente de Wikipedia en ingles
).
Vease tambien
[
editar
]
Referencias
[
editar
]
- ↑
Holtz, Thomas (2012).
Genus List for Holtz (2007) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages
. Consultado el 2 de noviembre de 2015
.
- ↑
Christine C. E. & Peterson, F. (2004). ≪Reconstruction of the Upper Jurassic Morrison Formation extinct ecosystem?a synthesis.≫
Sedimentary Geology
167
, 309-355.
- ↑
Liddell & Scott (1980).
Greek-English Lexicon, Abridged Edition
.
Oxford University Press
, Oxford, UK.
ISBN
0-19-910207-4
.
- ↑
Holland, W. J. (1913).
To the River Plate and Back. The narrative of a scientific mission to South America, with observations on things seen and suggested
. New York & London: G. P. Putnam's Sons.
- ↑
a
b
c
Lambert D. (1993)
The Ultimate Dinosaur Book
ISBN 0-86438-417-3
- ↑
a
b
Hartman, Scott (22 de julio de 2014).
≪Smackdown: Supersaurus vs. Giraffatitan and Diplodocus≫
(en ingles)
. Consultado el 5 de septiembre de 2015
.
- ↑
Upchurch, P., Barrett, P. M., Dodson, P. (2004).
≪Sauropoda≫
. En Weishampel, D. B. Dodson P, Osmolska, H., ed.
The Dinosauria (2nd Edition)
.
University of California Press
. pp.
316
.
ISBN
0-520-24209-2
.
- ↑
a
b
Paul, Gregory S. (2010).
Princeton Field Guide to Dinosaurs
. Princeton University Press.
ISBN
978-0-691-13720-9
.
- ↑
Paul, Gregory S. (Fall de 1994).
≪Big Sauropods - Really, Really Big Sauropods≫
.
The Dinosaur Report
. The Dinosaur Society. pp. 12-13.
- ↑
a
b
Foster, J.R. (2003). Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A. New Mexico Museum of Natural History and Science:Albuquerque, New Mexico. Bulletin 23.
- ↑
Mazzetta, Gerardo V.; Christiansen, Per; Farina, Richard A. (2004).
≪Giants and Bizarres: Body Size of Some Southern South American Cretaceous Dinosaurs≫
(PDF)
.
Historical Biology
16
(2-4): 71-83.
doi
:
10.1080/08912960410001715132
. Consultado el 8 de enero de 2008
.
- ↑
Seebacher, Frank (2001).
≪A new method to calculate allometric length-mass relationships of dinosaurs≫
.
The Society of Vertebrate Paleontology
21
(1): 51-60.
doi
:
10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2
. Consultado el 5 de septiembre de 2015
.
- ↑
a
b
Dodson, P.; Behrensmeyer, A.K.; Bakker, R.T.; McIntosh, J.S. (1980). ≪Taphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison Formation≫.
Paleobiology
6
: 208-232.
JSTOR
240035
.
- ↑
Coe, M.J.; Dilcher, D.L.; Farlow, J.O.; Jarzen, D.M.; Russell, D.A. (1987). ≪Dinosaurs and land plants≫. En Friis, E.M.; Chaloner, W.G.; Crane, P.R., eds.
The Origins of Angiosperms and Their Biological Consequences
. Cambridge University Press. pp. 225-258.
ISBN
0-521-32357-6
.
- ↑
Benson, R. B. J.; Campione, N. S. E.; Carrano, M. T.; Mannion, P. D.; Sullivan, C.; Upchurch, P.; Evans, D. C. (2014).
≪Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage≫
.
PLoS Biology
12
(5): e1001853.
PMC
4011683
.
PMID
24802911
.
doi
:
10.1371/journal.pbio.1001853
.
- ↑
Wedel, M. J. y Cifelli, R. L.
≪
Sauroposeidon
: Oklahoma’s Native Giant.≫
2005.
Oklahoma Geology Notes
65:2.
- ↑
a
b
Gillette, D.D., 1994,
Seismosaurus: The Earth Shaker
. New York, Columbia University Press, 205 pp
- ↑
a
b
Carpenter, K. (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod
Amphicoelias fragillimus
." In Foster, J.R. and Lucas, S.G., eds., 2006,
Paleontology and Geology of the Upper Jurassic Morrison Formation.
New Mexico Museum of Natural History and Science Bulletin
36
: 131?138.
- ↑
≪The biggest of the big≫
. Skeletaldrawing.com. 14 de junio de 2013
. Consultado el 26 de mayo de 2016
.
- ↑
a
b
Stevens, K.A.; Parrish, J.M. (1999). ≪Neck posture and feeding habits of two Jurassic sauropod dinosaurs≫.
Science
284
(5415): 798-800.
Bibcode
:
1999Sci...284..798S
.
PMID
10221910
.
doi
:
10.1126/science.284.5415.798
.
- ↑
a
b
c
d
e
Upchurch, P. & Barrett, P. M. (2000). Chapter 4: ≪The evolution of sauropod feeding mechanism.≫ En:
Evolution of Herbivory in Terrestrial Vertebrates
ISBN 0-521-59449-9
- ↑
Bakker, Robert T. (1986)
The Dinosaur Heresies: New Theories Unlocking the Mystery of the Dinosaurs and their Extinction
. New York: Morrow.
- ↑
Knoll, F., Galton, P. M., Lopez-Antonanzas, R. (2006). ≪Paleoneurological evidence against a proboscis in the sauropod dinosaur
Diplodocus
.≫
Geobios
,
39
: 215-221.
- ↑
Lawrence M. Witmer
et al.
, ≪Nostril Position in Dinosaurs and other Vertebrates and its Significance for Nasal Function.≫
Science
293
, 850 (2001).
- ↑
a
b
Wilson JA (2005). ≪Overview of Sauropod Phylogeny and Evolution≫. En Rogers, K. A. & Wilson, J. A. (eds), ed.
The Sauropods: Evolution and Paleobiology
. Indiana University Press. p. 15-49.
ISBN
0-520-24623-3
.
- ↑
Holland, W. J. (1915). ≪Heads and Tails: a few notes relating to the structure of sauropod dinosaurs.≫.
Annals of the Carnegie Museum
9
: 273-278.
- ↑
Myhrvold, N. P. y Currie, P. J. (1997).
≪Supersonic sauropods? Tail dynamics in the diplodocids≫
.
Paleobiology
23
: 393-409.
- ↑
a
b
Benton, Michael J. (2012).
Prehistoric Life
. Dorling Kindersley. pp.
268
?269.
ISBN
978-0-7566-9910-9
.
- ↑
Bonnan, M. F. (2003). ≪The evolution of manus shape in sauropod dinosaurs: implications for functional morphology, forelimb orientation, and phylogeny.≫
Journal of Vertebrate Paleontology
,
23
: 595-613.
- ↑
a
b
Czerkas, S. A. (1993). ≪Discovery of dermal spines reveals a new look for sauropod dinosaurs≫.
Geology
20
: 1068-1070.
doi
:
10.1130/0091-7613(1992)020<1068:dodsra>2.3.co;2
.
- ↑
Czerkas, S. A. (1994). "The history and interpretation of sauropod skin impressions." In
Aspects of Sauropod Paleobiology
(M. G. Lockley, V. F. dos Santos, C. A. Meyer, and A. P. Hunt, Eds.),
Gaia
No. 10
. (Lisbon, Portugal).
- ↑
a
b
Haines, T., James, J.
Time of the Titans
Archivado
el 31 de octubre de 2013 en
Wayback Machine
.
. ABC Online.
- ↑
a
b
c
d
e
f
Tschopp, E.; Mateus, O. V.; Benson, R. B. J. (2015).
≪A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda)≫
.
PeerJ
3
: e857.
PMC
4393826
.
PMID
25870766
.
doi
:
10.7717/peerj.857
.
- ↑
Tschopp, E.; Mateus, O. V. (2012). ≪The skull and neck of a new flagellicaudatan sauropod from the Morrison Formation and its implication for the evolution and ontogeny of diplodocid dinosaurs≫.
Journal of Systematic Palaeontology
11
(7): 1.
doi
:
10.1080/14772019.2012.746589
.
- ↑
Marsh, O. C.: ≪Principal characters of American Jurassic dinosaurs. Part I.≫
American Journal of Science
3; 411-416 (1878).
- ↑
McIntosh (2005).
≪The Genus Barosaurus (Marsh)≫
. En Carpenter, Kenneth and Tidswell, Virginia (ed.), ed.
Thunder Lizards: The Sauropodomorph Dinosaurs
. Indiana University Press. pp.
38
-77.
ISBN
0-253-34542-1
.
- ↑
Upchurch P, Barrett PM, Dodson P (2004).
≪Sauropoda≫
. En Weishampel DB, Dodson P, Osmolska H, ed.
The Dinosauria (2nd Edition)
. University of California Press. p.
305
.
ISBN
0-520-24209-2
.
- ↑
a
b
Tschopp, E.; Mateus, O. (2016). ≪Diplodocus Marsh, 1878 (Dinosauria, Sauropoda): proposed designation of D. carnegii Hatcher, 1901 as the type species≫.
Bulletin of Zoological Nomenclature
73
(1): 17-24.
doi
:
10.21805/bzn.v73i1.a22
.
- ↑
a
b
Marsh, O. C. 1884. ≪Principal characters of American Jurassic dinosaurs. Part VII. On the Diplodocidae, a new family of the Sauropoda.≫
American Journal of Science
3: 160-168.
- ↑
Upchurch, P., Barrett, P. M., y Dodson, P. (2004). "≪Sauropoda.≫ En D. B. Weishampel, H. Osmolska, y P. Dodson (eds.),
The Dinosauria (2nd edition)
. University of California Press, Berkeley 259-322.
- ↑
Brezinski, D. K.; Kollar, A. D. (2008). ≪Geology of the Carnegie Museum Dinosaur Quarry Site of Diplodocus carnegii, Sheep Creek, Wyoming≫.
Annals of Carnegie Museum
77
(2): 243-252.
doi
:
10.2992/0097-4463-77.2.243
.
- ↑
J. B. Hatcher. 1901. Diplodocus Marsh; its osteology, taxonomy, and probate habits, with a restoration of the skeleton. Memoirs of the Carnegie Museum 1:1-63
- ↑
Holland WJ. The skull of
Diplodocus
. Memoirs of the Carnegie Museum IX; 379?403 (1924).
- ↑
Gillette, D.D. (1991). ≪
Seismosaurus halli
, gen. et sp. nov., a new sauropod dinosaur from the Morrison Formation (Upper Jurassic/Lower Cretaceous) of New Mexico, USA≫.
Journal of Vertebrate Paleontology
11
(4): 417-433.
doi
:
10.1080/02724634.1991.10011413
.
- ↑
Lucas S, Herne M, Heckert A, Hunt A, and Sullivan R.
Reappraisal of
Seismosaurus
, A Late Jurassic Sauropod Dinosaur from New Mexico.
Archivado
el 8 de octubre de 2019 en
Wayback Machine
. The Geological Society of America, 2004 Denver Annual Meeting (7?10 November 2004). Retrieved on 2007-05-24.
- ↑
Lucas, S.G.; Spielman, J.A.; Rinehart, L.A.; Heckert, A.B.; Herne, M.C.; Hunt, A.P.; Foster, J.R.; Sullivan, R.M. (2006). ≪Taxonomic status of
Seismosaurus hallorum
, a Late Jurassic sauropod dinosaur from New Mexico≫. En Foster, J.R.; Lucas, S.G., eds.
Paleontology and Geology of the Upper Morrison Formation
. New Mexico Museum of Natural History and Science (bulletin 36). pp. 149-161.
ISSN
1524-4156
.
- ↑
Lovelace, David M.; Hartman, Scott A.; Wahl, William R. (2007). ≪Morphology of a specimen of
Supersaurus
(Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny≫.
Arquivos do Museu Nacional
65
(4): 527-544.
- ↑
a
b
Taylor, M.P. & Naish, D. 2005. ≪The phylogenetic taxonomy of Diplodocoidea (Dinosauria: Sauropoda).≫
PaleoBios
25(2): 1-7.
[1]
- ↑
a
b
Harris, J. D. 2006. ≪The significance of
Suuwassea emiliae
(Dinosauria: Sauropoda) for flagellicaudatan intrarelationships and evolution.≫
Journal of Systematic Palaeontology
4(2): 185-198.
- ↑
Bonaparte, J. F. & Mateus, O. 1999. ≪A new diplodocid,
Dinheirosaurus lourinhanensis
gen. et sp. nov., from the Late Jurassic beds of Portugal.≫
Revista del Museo Argentino de Ciencias Naturales
. 5(2):13-29.
[2]
Archivado
el 19 de febrero de 2012 en
Wayback Machine
.
- ↑
a
b
Rauhut, O. W. M., Remes, K., Fechner, R., Cladera, G., & Puerta, P. 2005. ≪Discovery of a short-necked sauropod dinosaur from the Late Jurassic period of Patagonia.≫
Nature
435: 670-672.
- ↑
a
b
Wilson, J. A., 2002, ≪Sauropod dinosaur phylogeny: critique and cladistica analysis≫.
Zoological Journal of the Linnean Society
136: 217-276.
- ↑
Wilson JA. 2002. Sauropod dinosaur phylogeny: critique and cladistic analysis. Zoological Journal of the Linnean Society 136: 217?276.
- ↑
Upchurch P, Barrett, P. M., Dodson P. (2004).
≪Sauropoda≫
. En Weishampel DB, Osmolska H, Dodson P, ed.
The Dinosauria (2nd Edition)
. University of California Press. pp.
316
.
ISBN
0-520-24209-2
.
- ↑
Whitlock, J. A. (2011). ≪A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda).≫
Zoological Journal of the Linnean Society
. Publicado en linea el 12 de enero de 2011.
- ↑
Schmitz, L.; Motani, R. (2011). ≪Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology≫.
Science
332
(6030): 705-8.
Bibcode
:
2011Sci...332..705S
.
PMID
21493820
.
doi
:
10.1126/science.1200043
.
- ↑
Hatcher JB. "
Diplodocus
(Marsh): Its osteology, taxonomy, and probable habits, with a restoration of the skeleton". Memoirs of the Carnegie Museum, vol. 1 (1901), pp. 1?63
- ↑
Kermack, Kenneth A. (1951). ≪A note on the habits of sauropods≫.
Annals and Magazine of Natural History
12
(4): 830-832.
- ↑
a
b
Gangewere, J.R. (1999). "
Diplodocus carnegii
Archivado
el 12 de enero de 2012 en
Wayback Machine
.". Carnegie Magazine.
- ↑
Pierson, D. J. (2009). ≪The Physiology of Dinosaurs: Circulatory and Respiratory Function in the Largest Animals Ever to Walk the Earth≫.
Respiratory Care
54
(7): 887-911.
PMID
19558740
.
doi
:
10.4187/002013209793800286
.
- ↑
Hay, Oliver P., ≪On the Habits and Pose of the Sauropod Dinosaurs, especially of Diplodocus.≫
The American Naturalist
, Vol. XLII, Oct. 1908.
- ↑
Holland, W. J., ≪A Review of Some Recent Criticisms of the Restorations of Sauropod Dinosaurs Existing in the Museums of the United States, with Special Reference to that of Diplodocus carnegiei in the Carnegie Museum.≫
The American Naturalist
, 44:259-283. 1910.
- ↑
Hay, O. P., 1910,
Proceedings of the Washington Academy of Sciences
, vol. 12, pp. 1-25.
- ↑
a
b
c
Stevens KA, Parrish JM (2005).
≪Neck Posture, Dentition and Feeding Strategies in Jurassic Sauropod Dinosaurs≫
. En Carpenter, Kenneth and Tidswell, Virginia (ed.), ed.
Thunder Lizards: The Sauropodomorph Dinosaurs
. Indiana University Press. pp.
212
-232.
ISBN
0-253-34542-1
.
- ↑
Upchurch, P,
et al.
(2000).
≪Neck Posture of Sauropod Dinosaurs≫
.
Science
287, 547b (2000);DOI: 10.1126/science.287.5453.547b
. Consultado el 28 de noviembre de 2006
.
- ↑
Norman, D.B. (1985).
The Illustrated Encyclopedia of Dinosaurs
. London: Salamander Books Ltd.
- ↑
Dodson, P. (1990). ≪Sauropod paleoecology.≫ En:
The Dinosauria
1st Edition, (Eds. Weishampel, D. B., Dodson, P. & Osmolska, H.)
- ↑
Barrett, P.M. & Upchurch, P. (1994). ≪Feeding mechanisms of
Diplodocus
.≫
Gaia
10
, 195-204.
- ↑
Henderson, Donald M. (2006).
≪Burly gaits: centers of mass, stability, and the trackways of sauropod dinosaurs≫
.
Journal of Vertebrate Paleontology
26
(4): 907-921.
- ↑
a
b
Mallison, H. (2011). ≪Rearing Giants ? kinetic-dynamic modeling of sauropod bipedal and tripodal poses≫. En Farlow, J.; Klein, N.; Remes, K.; Gee, C.; Snader, M., eds.
Biology of the Sauropod Dinosaurs: Understanding the life of giants. Life of the Past
. Indiana University Press.
ISBN
978-0-253-35508-9
.
- ↑
Paul, Gregory S. (2017). ≪Restoring Maximum Vertical Browsing Reach in Sauropod Dinosaurs≫.
The Anatomical Record
300
(10): 1802-1825.
doi
:
10.1002/ar.23617
.
- ↑
Barrett, P. M. & Upchurch, P. (2005). ≪Sauropodomorph Diversity through Time, Paleoecological and Macroevolutionary Implications.≫ En:
The Sauropods: Evolution and Paleobiology
(Eds. Curry, K. C.).
- ↑
Cobley, Matthew J.; Rayfield, Emily J.; Barrett, Paul M. (2013).
≪Inter-Vertebral Flexibility of the Ostrich Neck: Implications for Estimating Sauropod Neck Flexibility≫
.
PLoS ONE
8
(8): e72187.
Bibcode
:
2013PLoSO...872187C
.
PMC
3743800
.
PMID
23967284
.
doi
:
10.1371/journal.pone.0072187
.
- ↑
Ghose, Tia (15 de agosto de 2013).
≪Ouch! Long-Necked Dinosaurs Had Stiff Necks≫
. livescience.com
. Consultado el 31 de enero de 2015
.
- ↑
Taylor, M.P.
(2014).
≪Quantifying the effect of intervertebral cartilage on neutral posture in the necks of sauropod dinosaurs≫
.
PeerJ
2
: e712.
PMC
4277489
.
PMID
25551027
.
doi
:
10.7717/peerj.712
.
- ↑
Young, Mark T.; Rayfield, Emily J.; Holliday, Casey M.; Witmer, Lawrence M.; Button, David J.; Upchurch, Paul; Barrett, Paul M. (August 2012). ≪Cranial biomechanics of Diplodocus (Dinosauria, Sauropoda): testing hypotheses of feeding behaviour in an extinct megaherbivore≫.
Naturwissenschaften
99
(8): 637-643.
Bibcode
:
2012NW.....99..637Y
.
ISSN
1432-1904
.
PMID
22790834
.
doi
:
10.1007/s00114-012-0944-y
.
- ↑
D’Emic, M. D.; Whitlock, J. A.; Smith, K. M.; Fisher, D. C.; Wilson, J. A. (2013).
≪Evolution of high tooth replacement rates in sauropod dinosaurs≫
. En Evans, A. R., ed.
PLoS ONE
8
(7): e69235.
Bibcode
:
2013PLoSO...869235D
.
PMC
3714237
.
PMID
23874921
.
doi
:
10.1371/journal.pone.0069235
.
- ↑
Whitlock, John A.; Wilson, Jeffrey A.; Lamanna, Matthew C. (March 2010). ≪Description of a Nearly Complete Juvenile Skull of
Diplodocus
(Sauropoda: Diplodocoidea) from the Late Jurassic of North America≫.
Journal of Vertebrate Paleontology
30
(2): 442-457.
doi
:
10.1080/02724631003617647
.
- ↑
Moratalla, J. (2008).
Dinosaurios. Un paseo entre gigantes
. EDAF.
ISBN
978-84-414-2450-0
.
- ↑
Senter, P. (2006). ≪Necks for Sex: Sexual Selection as an Explanation for Sauropod Neck Elongation≫.
Journal of Zoology
271
(1): 45-53.
doi
:
10.1111/j.1469-7998.2006.00197.x
.
- ↑
Taylor, M.P.; Hone, D.W.E.; Wedel, M.J.; Naish, D. (2011).
≪The long necks of sauropods did not evolve primarily through sexual selection≫
.
Journal of Zoology
285
(2): 151-160.
doi
:
10.1111/j.1469-7998.2011.00824.x
.
- ↑
Chiappe, Luis y Lowell Dingus, 2001:
Walking on Eggs: The Astonishing Discovery of Thousands of Dinosaur Eggs in the Badlands of Patagonia
, Scribner.
- ↑
Grellet-Tinner, Chiappe, & Coria, ≪Eggs of titanosaurid sauropods from the Upper Cretaceous of Auca Mahuevo (Argentina).≫
Can. J. Earth Sci.
41(8): 949-960 (2004).
- ↑
Ruxton, Graeme D.; Birchard, Geoffrey F.; Deeming, D Charles (2014). ≪Incubation time as an important influence on egg production and distribution into clutches for sauropod dinosaurs≫.
Paleobiology
40
(3): 323-330.
doi
:
10.1666/13028
.
- ↑
Sander, P. M. (2000). ≪Long bone histology of the Tendaguru sauropods: Implications for growth and biology.≫
Paleobiology
26
, 466-488.
- ↑
Sander, P. M., N. Klein, E. Buffetaut, G. Cuny, V. Suteethorn, y J. Le Loeuff (2004). ≪Adaptive radiation in sauropod dinosaurs: Bone histology indicates rapid evolution of giant body size through acceleration.≫
Organisms, Diversity & Evolution
4
, 165-173.
- ↑
Sander, P. M., and N. Klein (2005). ≪Developmental plasticity in the life history of a prosauropod dinosaur.≫
Science
310
1800-1802.
- ↑
Trujillo, K.C.; Chamberlain, K.R.; Strickland, A. (2006). ≪Oxfordian U/Pb ages from SHRIMP analysis for the Upper Jurassic Morrison Formation of southeastern Wyoming with implications for biostratigraphic correlations≫.
Geological Society of America Abstracts with Programs
38
(6): 7.
- ↑
Bilbey, S.A. (1998). ≪Cleveland-Lloyd Dinosaur Quarry ? age, stratigraphy and depositional environments≫. En Carpenter, K.; Chure, D.; Kirkland, J.I., eds.
The Morrison Formation: An Interdisciplinary Study
. Modern Geology
22
. Taylor and Francis Group. pp. 87-120.
ISSN
0026-7775
.
- ↑
Russell, Dale A.
(1989).
An Odyssey in Time: Dinosaurs of North America
. Minocqua, Wisconsin: NorthWord Press. pp. 64-70.
ISBN
978-1-55971-038-1
.
- ↑
a
b
Mateus, Octavio (2006). ≪Jurassic dinosaurs from the Morrison Formation (USA), the Lourinha and Alcobaca Formations (Portugal), and the Tendaguru Beds (Tanzania): A comparison≫. En Foster, John R.; Lucas, Spencer G., eds.
Paleontology and Geology of the Upper Jurassic Morrison Formation
. New Mexico Museum of Natural History and Science Bulletin,
36
. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 223-231.
- ↑
Foster, J. (2007). "Appendix." Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. pp. 327?329.
- ↑
Chure, Daniel J.; Litwin, Ron; Hasiotis, Stephen T.; Evanoff, Emmett; Carpenter, Kenneth (2006). ≪The fauna and flora of the Morrison Formation: 2006≫. En Foster, John R.; Lucas, Spencer G., eds.
Paleontology and Geology of the Upper Jurassic Morrison Formation
. New Mexico Museum of Natural History and Science Bulletin,
36
. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 233-248.
- ↑
Foster, John R. (2003).
Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A
. New Mexico Museum of Natural History and Science Bulletin,
23
. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. p. 29.
- ↑
Carpenter, Kenneth
(2006). ≪Biggest of the big: a critical re-evaluation of the mega-sauropod
Amphicoelias fragillimus
≫. En Foster, John R.; Lucas, Spencer G., eds.
Paleontology and Geology of the Upper Jurassic Morrison Formation
. New Mexico Museum of Natural History and Science Bulletin,
36
. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 131-138.
- ↑
Bakker, Robert T.
(1986).
The Dinosaur Heresies: New Theories Unlocking The Mystery of the Dinosaurs and Their Extinction
. New York: William Morrow. p. 203.
ISBN
0140100555
.
- ↑
Mediateca (25 de noviembre de 2013).
≪Diplodocus carnegii: 100 anos en el Museo Nacional de Ciencias Naturales (1913-2013)≫
.
Museo Naciona de Ciencias Naturales. Comunicacion
. Consultado el 30 de septiembre de 2023
.
- ↑
Perez Garcia, A. y Sanchez Chillon, B. (2009). ≪Historia de
Diplodocus carnegii
del MNCN: primer esqueleto de dinosaurio montado en la Peninsula Iberica≫.
Revista Espanola de Paleontologia
24
(2): 133-148.
Enlaces externos
[
editar
]
 Portal:Dinosaurios
. Contenido relacionado con
Dinosaurios
.
Portal:Dinosaurios
. Contenido relacionado con
Dinosaurios
.
 Wikimedia Commons
alberga una galeria multimedia sobre
Diplodocus
.
Wikimedia Commons
alberga una galeria multimedia sobre
Diplodocus
.
 Wikispecies
tiene un articulo sobre
Diplodocus
.
Wikispecies
tiene un articulo sobre
Diplodocus
.
- Diplodocus
in the Dino Directory
(en ingles)
- Museo Nacional de Ciencias Naturales, Madrid
- Diplodocus Marsh, by J.B. Hatcher 1901
- Su Osteologia, Taxonomia, y Probables Habitos, con una Reconstruccion del Esqueleto.
Memorias del Museo Carnegie
, Vol. 1, N.º 1, 1901. Texto completo, de lectura libre
(en ingles)
.
- Museo Carnegie de Historia Natural - Historia
(en ingles)
- Reconstrucciones esqueleticas de diplodocidos
incluyendo
D. carnegii
,
D. longus
, y
D. hallorum
, de Scott Hartman's Skeletal Drawing website
(en ingles)
.
- Diplodoco en duiops
- Diplodoco en Dinodata
(en ingles)