Unter
Evolution
(von
lateinisch
evolvere
?herausrollen“, ?auswickeln“, ?entwickeln“) versteht man im deutschsprachigen Raum in erster Linie die
biologische Evolution
. Darunter wird die von
Generation
zu Generation stattfindende allmahliche Veranderung der vererbbaren
Merkmale
einer
Population
von
Lebewesen
und von anderen organischen Strukturen (z. B.
Viren
) verstanden. Das Lehr- und Forschungsgebiet der Evolution wird als
Evolutionsbiologie
bezeichnet und unterliegt, wie viele andere Wissenschaften, einem kontinuierlichen Erkenntnisfortschritt. Hierzu konnen insbesondere neue Einsichten durch die Entdeckung neuer Fossilien oder die Anwendung neuer
Forschungsmethoden
beitragen. Das Themenfeld der Evolution wurde zuweilen unterteilt in die
Evolutionsgeschichte
, in der die Veranderungen der Lebewesen im Laufe der Erdgeschichte beschrieben werden und bei dem es Uberlappungen mit der
Palaontologie
gibt, sowie in die
Evolutionstheorie
, die naturwissenschaftliche Erklarungen (Hypothesen und Theorien) fur das Gesamtphanomen der Evolution entwickelt. Die beiden Ansatze sind heutzutage in der Wissenschaft innig miteinander verwoben und befruchten sich wechselseitig. Wissenschaftler beschaftigen sich ebenfalls im Rahmen der
theoretischen Biologie
mit der biologischen Evolution. Die theoretische Biologie als interdisziplinares Teilgebiet der Biologie entwickelt
mathematische Modelle
und fuhrt
statistische Hypothesentests
und
Laborexperimente
durch, um den Erkenntnisgewinn zu fordern.
Insbesondere seit der Begrundung der Evolutionstheorie hat der
Begriff der Evolution
eine erheblich Ausweitung in viele verschiedene Wissenschaftsbereiche erfahren, die zu einer großen Zahl recht unterschiedlicher Definitionen gefuhrt hat.
Geschichte
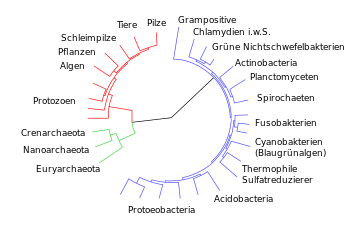 Stammbaum
-Darstellung der irdischen Organismen auf Basis molekularer Merkmale.
[1]
Die drei
Domanen
sind farblich getrennt dargestellt (blau:
Bakterien
; grun:
Archaeen
; rot:
Eukaryoten
).
Stammbaum
-Darstellung der irdischen Organismen auf Basis molekularer Merkmale.
[1]
Die drei
Domanen
sind farblich getrennt dargestellt (blau:
Bakterien
; grun:
Archaeen
; rot:
Eukaryoten
).
Die Merkmale der Lebewesen sind in Form von
Genen
codiert, die bei der
Fortpflanzung
kopiert und an die Nachkommen weitergegeben werden. Durch
Mutationen
entstehen unterschiedliche Varianten (
Allele
) dieser Gene, die zur Entstehung veranderter oder neuer Merkmale fuhren konnen. Diese Varianten sowie
Rekombinationen
fuhren zu erblich bedingten Unterschieden in Form der
genetischen Variabilitat
zwischen Individuen. Evolution findet statt, wenn sich die Haufigkeit bestimmter Allele in einer Population (die
Allelfrequenz
im
Genpool
) andert und die entsprechenden Merkmale in der Population dadurch seltener oder haufiger werden. Dies geschieht entweder durch
naturliche Selektion
(unterschiedliche Uberlebens- und Reproduktionsrate
aufgrund
dieser Merkmale), durch
sexuelle Selektion
oder zufallig durch
Gendrift
.
 Charles Darwin
im Alter von 51 Jahren, kurz nach der Veroffentlichung des Buches
On the Origin of Species
Charles Darwin
im Alter von 51 Jahren, kurz nach der Veroffentlichung des Buches
On the Origin of Species
 Alfred Russel Wallace
, etwa 1895
Alfred Russel Wallace
, etwa 1895
Der entscheidende Bruch mit dem fruheren Konzept konstanter typologischer Klassen oder Typen der Biologie kam mit der Theorie der Evolution durch naturliche Selektion, die von
Charles Darwin
und
Alfred Russel Wallace
in Form von im Laufe der Zeit veranderlichen Populationen formuliert wurde. Diese Bedeutung der Variabilitat und der durch naturliche Selektion in Gang gehaltene Prozess bei Lebewesen wurden dann erstmals ausfuhrlich und fundiert von Charles Darwin in seinem 1859 erschienenen Buch
The Origin of Species
dargestellt. Um 1900 wurden die Prinzipien der Vererbung bekannt (da die fruheren Untersuchungen von Gregor Mendel nicht weiter beachtet worden waren) und um 1910 die Bedeutung der Chromosomen. Dadurch schienen sich die Konzepte des
Darwinismus
, der die Veranderungen hervorhob, und der
Genetik
, die die statische Weitergabe von Merkmalen lehrte, einander zu widersprechen. Erst ab den 1930er Jahren konnten die Selektionsprozesse mit den
mendelschen Regeln zur Vererbung
in Einklang gebracht werden, woraus sich die
Synthetische Theorie der Evolution
entwickelte. Sie definierte Evolution als die zeitliche Anderung der relativen Allelhaufigkeiten (Allelfrequenzen) in einer Population.
[2]
Durch ihre
deskriptiven
und
kausalen
Aussagen wurde diese Theorie zum zentralen organisierenden Prinzip der modernen Biologie und lieferte eine fundierte Erklarung fur die Entstehung der Vielfalt des Lebens auf der Erde.
1944 lieferten die Arbeiten von
Oswald Avery
und seinen Kollegen ein starkes Indiz dafur, dass
DNA
der Trager genetischer Informationen ist, denn bislang hatte man eher Proteine ?im Verdacht“, entsprechende Informationen zu beherbergen. Zusammen mit der Entschlusselung der Struktur der DNA durch
Rosalind Franklin
,
James Watson
und
Francis Crick
im Jahr 1953 wurde die physische Basis der Vererbung geklart. Seitdem ist auch die molekulare Genetik ein zentraler Bestandteil der
Evolutionsbiologie
.
[3]
Grundlagen
Vererbung
Gregor Mendel zeigte anhand von Erbsenpflanzen, dass Vererbung in eng definierten (diskreten) Einheiten erfolgt, indem
Merkmale
von der Elterngeneration an die Nachkommen vererbt werden, und dass diese Merkmale diskret sind: Wenn ein Elternteil runde und der andere faltige Erbsen bildete, dann bildete der Nachwuchs nicht ein Gemisch, sondern entweder runde oder faltige Erbsen. Mendel wies außerdem nach, dass die Merkmale der Eltern in einer genau definierten und vorhersagbaren Weise an die Nachkommen vererbt wurden, namlich nach den
mendelschen Regeln
. Seine Forschungen waren die Basis fur das Konzept der diskreten, erblichen Merkmale, der
Gene
.
[4]
Mendels Arbeiten beantworteten die lange offene Frage, warum Merkmalsvarianten in Populationen stabil bleiben. Im Nachhinein muss man feststellen, dass es ein großer Zufall war, dass er lauter diskrete Merkmale gewahlt hatte, denn bei vielen anderen Merkmalen (etwa bezuglich der erreichten Pflanzenhohe) waren komplexere genetische Einflusse (siehe
Polygenie
) und auch umweltbedingte Einflusse (siehe
Modifikation)
wirksam geworden.
Spatere Forschungen enthullten die physische Basis der Gene und identifizierten die
DNA
als das genetische Material. Gene wurden neu definiert als spezifische Regionen der DNA. DNA wird von Lebewesen als
Chromosomen
gelagert. Ein bestimmter Ort auf einem Chromosom wird als
Genlocus
(oder kurz
Locus
) bezeichnet, die Variante einer DNA-Sequenz auf einem bestimmten Locus bezeichnet man als
Allel
. Die Kopie der DNA erfolgt nicht perfekt und Anderungen (
Mutationen
) der Gene produzieren neue Allele und beeinflussen daher die Merkmale, die von diesen Genen kontrolliert werden. Diese einfache Beziehung zwischen einem Gen und einem Merkmal liegt in vielen Fallen vor, komplexe Merkmale, zum Beispiel die Widerstandsfahigkeit gegen Krankheiten, werden jedoch von vielen zusammenwirkenden Genen (?polygen“) kontrolliert.
[5]
 Die Struktur eines Ausschnittes einer DNA-
Doppelhelix
Die Struktur eines Ausschnittes einer DNA-
Doppelhelix
Genetische Variabilitat
Die genetische Variabilitat oder Variation resultiert aus Mutationen der DNA, der Wanderung von Individuen zwischen Populationen (dem
Genfluss
), und der Durchmischung von Genen bei der sexuellen Fortpflanzung (
Rekombination
). Bei einigen Lebensformen, wie Bakterien und Pflanzen, wird Variabilitat auch durch die Mischung des genetischen Materials zwischen Arten durch horizontalen Genfluss und
Hybridisierung
erzeugt.
[6]
[7]
Trotz all dieser Variabilitat verursachenden Prozesse sind die meisten Bereiche der DNA einer Art (das
Genom
) bei allen Individuen einer Art identisch.
[8]
Vergleichsweise kleine Anderungen des
Genotyps
(des Merkmale codierenden Teiles des Genoms), konnen jedoch erhebliche Auswirkungen auf den
Phanotyp
(die Gesamtheit der genetisch bedingten Merkmale eines Individuums; vereinfacht ausgedruckt, auf das genetisch bedingte außere Erscheinungsbild) haben. Zum Beispiel betragt der Unterschied der DNA-Sequenzen von
Schimpanse
und Mensch nur funf Prozent.
[9]
[10]
Der Phanotyp resultiert aus der Interaktion seiner individuellen genetischen Ausstattung, seines Genotyps, mit der Umwelt. Die Variabilitat der vererbbaren Merkmale innerhalb einer Population reflektiert also die Variabilitat des Genoms innerhalb dieser Population. Die Frequenz einzelner Merkmalsvarianten kann in einer Population schwanken und in Relation zu anderen Allelen des Gens großer oder kleiner werden. Alle evolutionar wirksamen Krafte agieren, indem sie diese Anderungen der Allelfrequenzen in die eine oder andere Richtung fordern. Die Variabilitat eines Merkmals verschwindet, wenn ein Allel eine feste Frequenz erreicht, wenn es also entweder aus der Population verschwindet oder wenn es alle anderen, fruher vorhandenen Allele ersetzt hat.
[11]
Homologie
Organe der Saugetiere wie Wirbelsaule, Augen, Verdauungskanal, Lungen ahneln denen anderer Wirbeltiere. Diese Gemeinsamkeiten basieren auf der Abstammung von gemeinsamen Vorfahren. Diese Ahnlichkeiten werden in der Biologie als Homologie bezeichnet. Fossile Skelettfunde bieten die Moglichkeit, Homologien zu erkennen und dadurch Hinweise auf eine gemeinsame Abstammung zu erlangen. Im Verlauf der Evolution haben viele Organismen ihre Lebensweise verandert. Durch genetische Variabilitat und naturliche Auslese fand ein Funktionswandel von Organen statt. Dadurch kam es zu einer Angepasstheit des Baues an die jeweilige Funktion.
Analogie
Analoge Organe weisen verschiedenen Grundbauplane auf, dennoch besitzen sie Ahnlichkeiten, die von ihrer Abstammung unabhangig sind. Der Grund dafur ist die Anpassung an gleiche Funktionen. Ein Beispiel dafur ist die Grabhand des
Maulwurfs
und die Grabschaufeln der
Maulwurfsgrille
, denn sie sind sich sehr ahnlich. Wahrend die Hand des Maulwurfs ein Knochenskelett aufweist, liegt bei der Maulwurfsgrille ein Außenskelett aus
Chitin
vor. Maulwurf und Maulwurfsgrille besitzen unterschiedliche Bauplane ihrer außerlich ahnlichen Gliedmaßen. Analoge Ahnlichkeiten sind stammesgeschichtlich unabhangig voneinander, lassen jedoch Ruckschlusse auf ahnliche Umweltbedingungen und Lebensweisen zu.
Evolutionsfaktoren
Als Evolutionsfaktoren bezeichnet man in der Biologie Prozesse, durch die der
Genpool
(die Gesamtheit aller Genvariationen in einer
Population
) verandert wird. Dies erfolgt uberwiegend durch Veranderungen der
Allelfrequenzen
im Genpool der Population. Diese Prozesse sind die zentrale Ursache fur evolutionare Veranderungen.
Die wesentlichen Evolutionsfaktoren, die den Genpool (Gesamtheit aller Genvarianten in einer Population) verandern, sind
Mutation
,
Rekombination
,
Selektion
und
Gendrift
.
Mutation
 Duplikation eines Abschnittes eines
Chromosoms
Duplikation eines Abschnittes eines
Chromosoms
Mutationen konnen im Korper an irgendeiner Stelle außerhalb der
Keimbahn
, also außerhalb von Fortpflanzungszellen, auftreten und heißen dann somatische Mutation. Diese haben, im Gegensatz zu den Keimbahnmutationen, keinen direkten Einfluss auf die nachfolgenden Generationen, hochstens indirekt, wenn die somatische Mutation die Fitness des Tragerorganismus beeintrachtigt und dadurch die Weitergabewahrscheinlichkeit der eigenen Gene statistisch reduziert ist. Keimbahnmutationen sind Mutationen, die an die
Nachkommen
uber die Keimbahn
vererbt
werden konnen; sie betreffen
Eizellen
oder
Spermien
sowie deren Vorlaufer vor und wahrend der
Oogenese
bzw.
Spermatogenese
. Auf den Tragerorganismus, in dem sie stattfinden, haben sie normalerweise keinen Einfluss.
Mutationen und ihre Wirkungen sind feststellbar: So kann zum Beispiel die Entstehung neuartiger Enzyme in Mikroorganismen aufgrund der kurzeren Generationszeit im Zeitraffer beobachtet werden. Beispiele fur neu entstandene
Enzyme
sind die
Nylonasen
.
Rekombination
Rekombinationen sind Neuanordnungen von bestehenden Genen. Sie konnen im Rahmen der vermutlich phylogenetisch alteren parasexuellen Rekombination (bei Prokaryoten und einigen Pilzen) auftreten, aber auch im Rahmen der sexuellen Fortpflanzung. Bei letzterer, die typisch fur fast alle Pflanzen und Tiere ist, unterscheidet man die
Intrachromosomale Rekombination
durch Neukombination von
Allelen
innerhalb von Chromosomen (als Folge des
Crossing-overs
anlasslich der 1. Reifeteilung) und die
Interchromosomale Rekombination
durch Neukombination ganzer
Chromosomen
im
Chromosomensatz
.
Selektion
Selektion tritt auf, wenn Individuen mit fur das Uberleben und die Fortpflanzung vorteilhaften Merkmalen mehr Nachwuchs produzieren konnen als Individuen ohne diese Merkmale. Auf diese Weise konnen fur die Population im Laufe der Generationen insgesamt verbesserte Anpassungen an die Umweltbedingungen entstehen. Im Rahmen solcher Merkmalsanderungen kann sich eine Art auch im Rahmen der
Artbildung
in neue Arten aufspalten. Die ?normale“ Selektion lauft zwischen Individuen unterschiedlicher Spezies ab und wird auch als
naturliche Selektion
bezeichnet. Ein Sonderfall ist die durch den Menschen eingesetzte ?kunstliche Selektion“ oder
Zucht
, die beispielsweise fur die zahlreichen Hunderassen verantwortlich ist. Sobald Haushunde in der freien Wildbahn uberleben mussen, was in vielen Landern der Erde ein verbreitetes Phanomen ist, setzen sich durch naturliche Selektion bald nur bestimmte Genotypen durch. Die Hunde werden relativ einheitlich in der Große, in Farbcharakteristiken und im Verhalten, da sich die ?extremeren“ angezuchteten Eigenschaften in naturlicher Umgebung als nachteilig erweisen und die entsprechenden Hunde zu geringerem Fortpflanzungserfolg kommen lassen.
 Der Schwanz eines mannlichen
Pfaus
ist ein klassisches Beispiel fur ein sexuell selektiertes Merkmal.
Der Schwanz eines mannlichen
Pfaus
ist ein klassisches Beispiel fur ein sexuell selektiertes Merkmal.
Ein Spezialfall der Selektion bzw. der naturlichen Selektion ist die
sexuelle Selektion
, die intraspezifisch (also innerhalb einer Art) wirkt: Die Selektion auf Merkmale, deren Prasenz direkt mit dem Kopulationserfolg durch bevorzugte Partnerwahl korreliert ist.
[12]
Durch sexuelle Selektion evolvierte Merkmale sind besonders bei den Mannchen von Tieren verbreitet. Obwohl diese Merkmale die Uberlebenswahrscheinlichkeit einzelner Mannchen reduzieren konnen (z. B. durch behindernde Geweihe, durch Paarungsrufe oder leuchtende Farben), ist der Reproduktionserfolg solcher Mannchen im Normalfall hoher.
[13]
Helfersysteme und Eusozialitat stellen weitere Spezialfalle dar: Bei mehr als 200 Vogelarten und etwa 120 Saugerarten findet man soziale Strukturen, bei denen ein Teil der Individuen zumindest zeitweise auf eine eigene Reproduktion verzichtet und stattdessen Artgenossen bei deren Reproduktion unterstutzt. Dies steht in scheinbarem Widerspruch zu Darwins Thesen. Untersuchungen dieser Helfersysteme haben jedoch gezeigt, dass diese Hilfe meist umso starker erfolgt, je naher die Helfer mit dem aufzuziehenden Nachwuchs verwandt sind.
[14]
Da ein Teil des Genoms von Helfer und aufgezogenem Fremdnachwuchs identisch ist, erreicht der Helfer also trotz Verzicht auf eigene Reproduktion eine Weitergabe eines Teils seines Genoms. Da die Selektion hier nicht mehr auf der Ebene des Phanotyps, sondern des Genotyps ansetzt, hat
Richard Dawkins
fur diese und ahnliche Falle den Begriff des ?egoistischen Gens“ gepragt.
[15]
Bei
eusozialen
Insekten wie beispielsweise
Ameisen
und Sozialen
Faltenwespen
verzichtet der großte Teil der Weibchen lebenslang auf eine eigene Fortpflanzung. Eusozialitat ist fur diese Weibchen nicht mit einer verringerten evolutionaren Fitness verbunden, da sie aufgrund einer genetischen Besonderheit (
Haplodiploidie
) mit ihren Schwestern naher verwandt sind als mit potentiellen eigenen Nachkommen. Bei der Aufzucht von Schwestern geben sie also einen großeren Teil ihres Genoms weiter als bei der Aufzucht eigener Tochter.
[16]
Gendrift
Gendrift ist die Anderung von Allelfrequenzen von einer Generation zur nachsten, die geschieht, weil die Allele einer Generation von Nachkommen statistisch gesehen eine
Zufallsstichprobe
der Allele der Elterngeneration darstellen und deren Auswahl daher auch einem Zufallsfehler unterliegt. Selbst wenn keine Selektion stattfindet, tendieren Allelfrequenzen dazu, im Verlauf der Zeit großer oder kleiner zu werden, bis sie schließlich die Werte 0 % oder 100 % erreichen (?Fixierung“ des Allels). Schwankungen der Allelfrequenzen in aufeinanderfolgenden Generationen konnen daher durch reinen Zufall dazu fuhren, dass einzelne Allele aus der Population verschwinden. Zwei getrennte Populationen mit anfanglich gleichen Allelfrequenzen konnen daher durch zufallige Schwankungen in zwei unterschiedliche Populationen mit einem unterschiedlichen Satz von Allelen auseinanderdriften.
[17]
Ob naturliche Selektion oder Gendrift den großeren Einfluss auf das Schicksal neuer Mutationen haben, hangt von der Große der Population und der Starke der Selektion ab.
[18]
Naturliche Selektion dominiert in großen Populationen, Gendrift in kleinen. Schließlich hangt die Zeit, die ein Allel benotigt, um in einer Population durch Gendrift eine feste Frequenz zu erreichen (bis also 0 % oder 100 % der Individuen der Population das Allel tragen), von der Populationsgroße ab; bei kleineren Populationen geschieht dies schneller.
[19]
Die Große einer Population (genauer die
effektive Populationsgroße
) hat daher einen großen Einfluss auf den Verlauf der Evolution. Wenn eine Population beispielsweise durch einen
genetischen Flaschenhals
(eine vorubergehend sehr niedrige Populationsgroße) geht, verliert sie damit auch einen großen Teil ihrer genetischen Variabilitat. Die Population wird insgesamt gleichartiger und verliert viele seltene Varianten. Solche ?Flaschenhalse“ konnen durch Katastrophenereignisse, Klimaschwankungen, durch Wanderungen oder Teilung von Populationen sowie naturlich durch einen anthropogenen Belastungsdruck verursacht werden.
Entwicklung biologischer Komplexitat
Konzepte betreffend die Entwicklung biologischer Komplexitat
Ein wichtiges Ergebnis des Evolutionsprozesses ist die Entwicklung biologischer
Komplexitat
.
[20]
Die Evolution hat einige bemerkenswert komplexe Organismen hervorgebracht. Allerdings ist ein numerischer Grad der Komplexitat in der Biologie sehr schwer zu definieren oder zu messen. Als mogliche Messgroßen wurden beispielsweise Eigenschaften wie der Genomgroße, Anzahl der Gene, Anzahl der Zelltypen oder die Morphologie vorgeschlagen.
[21]
[22]
[23]
Normalerweise haben Organismen, die eine hohere Reproduktionsrate als ihre Konkurrenten haben, einen evolutionaren Vorteil. Um sich schneller vermehren und mehr Nachkommen produzieren zu konnen, konnen sich Organismen in Richtung großerer Einfachheit entwickeln, da sie dann weniger Ressourcen zur Fortpflanzung benotigen. Ein gutes Beispiel sind
Parasiten
wie der
Malaria
erreger
Plasmodium
und
Mykoplasmen
? diese Organismen verzichten auf Eigenschaften, die durch den Parasitismus uberflussig geworden sind.
[24]
Eine Abstammungslinie kann auch Komplexitat einbußen, wenn ein bestimmtes komplexes Merkmal in der
Umgebung
einfach keinen Selektionsvorteil bietet. Selbst wenn der Verlust dieses Merkmals keinen Selektionsvorteil bedeutet, kann durch das Merkmal durch eine Anhaufung von Mutationen verloren gehen, wenn diese neutral sind (keinen unmittelbaren Selektionsnachteil bedeuten).
[25]
 Passive und aktive Trends in der Komplexitat. Die Organismen im Anfangszustand sind rot dargestellt. Von einem aktiven Trend spricht man, wenn vereinfachende Mutationen einen selektiven Nachteil bedeuten. Bei einem passiven Trend ist das nicht der Fall, mit der Zeit entsteht zwar immer hohere Komplexitat, diese wird aber von immer weniger Arten getragen.
Passive und aktive Trends in der Komplexitat. Die Organismen im Anfangszustand sind rot dargestellt. Von einem aktiven Trend spricht man, wenn vereinfachende Mutationen einen selektiven Nachteil bedeuten. Bei einem passiven Trend ist das nicht der Fall, mit der Zeit entsteht zwar immer hohere Komplexitat, diese wird aber von immer weniger Arten getragen.
Gabe es in der Evolution (wie im 19. Jahrhundert weithin angenommen) einen aktiven Trend zur Komplexitat (
Orthogenese
),
[26]
dann waren zu erwarten, dass der haufigste Wert (der
Modus
) der Komplexitat unter den Organismen im Laufe der Zeit aktiv zunimmt.
[27]
Eine Zunahme der Komplexitat kann aber auch durch einen passiven Prozess erklart werden: Unter der Annahme,
- dass sich die Komplexitat rein zufallig andert (ohne im
teleologischen
Sinn eine bestimmte Richtung zu bevorzugen) und
- dass es eine Mindestkomplexitat gibt, nimmt die durchschnittliche Komplexitat der Biosphare im Laufe der Zeit zu. Dies beinhaltet eine Zunahme der
Varianz
, aber der Modus andert sich nicht.
[27]
Es gibt dann zwar auch eine Tendenz, dass im Laufe der Zeit einige Organismen mit immer hoherer Komplexitat entstehen, dies betrifft aber einen immer kleineren Prozentsatz der Lebewesen.
[23]
Einige wenige Lebewesen
entwickeln sich gelegentlich nach rechts und verlangern so den rechten ?Schwanz“ in der Komplexitats
verteilung
. Einige bewegen sich auch nach links, stoßen aber ganz links schließlich an eine Grenze, die durch Minimalanforderungen des Lebens (siehe etwa
Minimalgenom
) gegeben ist.
[28]
[29]
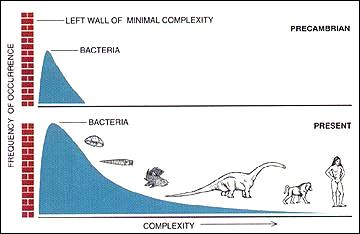 Haufigkeitsverteilung zellularer Organismen in Abhangigkeit von deren Komplexitat (im
Prakambrium
und heute). Die ?linke Wand der minimalen Komplexitat“ zeigt die durch das
Minimalgenom
gegebene untere Grenze an. Diese Darstellung erfolgt auf der Grundlage palaontologischer Beobachtungen.
Stephen J. Gould
,
[28]
[23]
ahnlich in der s/w-Ausgabe.
[29]
Haufigkeitsverteilung zellularer Organismen in Abhangigkeit von deren Komplexitat (im
Prakambrium
und heute). Die ?linke Wand der minimalen Komplexitat“ zeigt die durch das
Minimalgenom
gegebene untere Grenze an. Diese Darstellung erfolgt auf der Grundlage palaontologischer Beobachtungen.
Stephen J. Gould
,
[28]
[23]
ahnlich in der s/w-Ausgabe.
[29]
Im passiven Fall ist jeder Anschein einer Evolution, die von sich aus (aktiv/teleologisch) zu immer komplexeren Organismen fuhrt, darauf zuruckzufuhren, dass sich die Wahrnehmung des Menschen auf die wenigen großen, komplexen Organismen am rechten Ende der Komplexitatsverteilung (den Menschen selbst und andere ?hohere“ Tiere und Pflanzen) konzentrieren und einfachere und viel haufigere Organismen ignorieren (vgl.
Mikrobielle Dunkle Materie
) ? außer naturlich in der Fruhzeit der Erde, als es nur Lebewesen mit einfacher Organisation gab. Das passive Modell sagt voraus, dass die Mehrheit der Arten mikroskopische
Prokaryoten
sind, was durch Schatzungen von 10
6
bis 10
9
rezenten
Prokaryotenspezies
[30]
im Vergleich zu Diversitatsschatzungen von 10
6
bis 3·10
6
fur die
Eukaryoten
(komplex-zellularen Organismen:
Protisten
,
Pflanzen
,
Pilz
und
Tiere
? inkl. dem Menschen)
[31]
[32]
bestatigt wird.
Die Prokaryoten (
Bakterien
und
Archaeen
) sind bis heute am haufigsten und erfolgreichsten geblieben, und der Modus hat seine bei diesen Organismen gelegene Position nicht mehr verandert.
[28]
[29]
[23]
Die von
Stephen J. Gould
et al.
zunachst nur fur
zellulare Organismen
dargestellte Verteilung lasst sich im Ubrigen auf der linken Seite noch ein Stuck fortsetzen, indem man auch
Viren
in Betracht zieht. Die minimale Organisationsgroße (?linke Mauer“) der Viren ist noch um einiges geringer als die der zellularen Organismen. Dies geschieht aber unter Preisgabe der Unabhangigkeit von
Wirtsorganismen
. Wichtig ist, dass die zu erwartende Vielfalt in diesem Bereich nochmals großer ist als bei den Prokaryoten. Der Modus der Verteilung verschiebt sich weiter nach links.
[33]
Evolution des Bewusstseins
 Ist das ?Bild der Welt im Kopf“ evolutionar erklarbar?
Ist das ?Bild der Welt im Kopf“ evolutionar erklarbar?
Dass das
Bewusstsein
ebenfalls im Laufe der
Evolution des Menschen
als Folge hochkomplexer
neuronaler Vernetzung
entstanden ist, wird von den Naturwissenschaften heute allgemein angenommen. Bislang ungeklart ist die Frage, wie das zu erklaren ist. Vom Standpunkt des herrschenden
physikalistischen
Paradigmas
mussten die
physikalischen Gesetze
fundamental sein, sodass chemische, biologische und selbst
geistige
Vorgange
(
Wahrnehmung
, Bewusstsein,
Emotionen
,
Kognition
etc.)
auch als
abhangige Eigenschaften
des Physischen betrachtet werden konnen
(?sie
supervenieren
auf der Physis“)
.
[34]
Extrempositionen wie der
Epiphanomenalismus
oder der
Substanzdualismus
werden heute kaum noch vertreten. Erstere degradiert den Geist auf eine bloße, ganzlich uneigenstandige Begleiterscheinung; zweitere setzt Geist als vom Korperlichen unabhangige ?Substanz“ voraus. Es wird zumeist anerkannt, dass geistige Prozesse sich fundamental von physischen Prozessen unterscheiden: Das ?
innere Abbild der Welt
“ und die damit verbundenen
Erlebniseindrucke
konnen nicht bewiesen werden, denn sie sind nicht direkt beobachtbar, nicht mess- oder berechenbar, lassen sich nicht auf zugehorige
Hirnstrommuster
reduzieren und weisen zum Teil Eigenschaften auf, die den Naturgesetzen zu widersprechen scheinen.
[35]
[36]
Damit kann daruber im Wesentlichen nur noch
philosophiert
werden.
[37]
In der
Philosophie des Geistes
wird die Verbindung geistiger und korperlicher Prozesse seit langem kontrovers diskutiert. Doch auch etliche Biologen ? allen voran
Ernst Mayr
? wiesen darauf hin, dass Prozesse des Lebens oder der Evolution nicht auf die Gesetze der
Thermodynamik
reduziert werden konnten, sondern uberdies unabhangigen, originar biologischen Mechanismen unterlagen.
[38]
Die meisten gegenwartigen Erklarungsversuche sind
eigenschaftsdualistisch
, das heißt, Physis
und
Psyche werden als zwei getrennte Eigenschaften
einer
?Substanz“ aufgefasst. Strittig ist jedoch vor allem, ob nur das Mentale vom Korper abhangig ist oder ob auch umgekehrt ein Einfluss besteht ? das Mentale also gewissermaßen ?gleichwertig“ ist. Die alltagliche Erfahrung, dass unser Bewusstsein unser Handeln vielfach
frei
bestimmt ? die so genannte
Mentale Verursachung
? ist das großte Problem der Debatte.
[39]
Die gangigste Erklarung fur die evolutionare Entstehung von Geist wird als
nicht-reduktiver Physikalimsus
bezeichnet: Die Vertreter solcher Theorien gehen davon aus, dass Geist irgendwann als
emergentes Phanomen
biochemischer Prozesse ab einem bestimmten Komplexitatsgrad vernetzter Zellen spontan auftauchte (daher auch
Moderner Emergentismus
):
Emergenz
ist die Herausbildung von neuen
Systemeigenschaften
infolge des (
holistischen
) Zusammenspiels seiner Elemente. Solche ganzlich neuen Phanomene lassen sich nicht aus der (
reduktionistischen
) Untersuchung der einzelnen Elemente herleiten.
[40]
Fur die
mentale Verursachung
werden vor allem
funktionalistische
Losungsansatze favorisiert.
[41]
Kritiker verweisen darauf, dass die extreme Andersartigkeit des Geistigen eine ?starke Emergenz“ voraussetzen wurde,
[42]
was einem
Wunder
gleichkame.
[43]
[44]
Bereits
Ernst Haeckel
[40]
im ausgehenden 19. Jahrhundert, der Evolutionsbiologe
Bernhard Rensch
[45]
in der Mitte des 20. Jahrhunderts sowie einige Philosophen ? unter ihnen
Alfred North Whitehead
(ab 1919),
[40]
Pierre Teilhard de Chardin
(1955) und (auf beiden aufbauend)
Patrick Spat
(2010) ? vertreten daher
panpsychistische
Positionen, denen die Annahme gemeinsam ist, dass
Geist
? im Sinne einer ?immateriellen Innerlichkeit, Fahigkeit oder Kraft“ ? von Anfang an in jeglicher Materie vorhanden ist. Deutliche Hinweise darauf werden in der Existenz ?ungewohnlicher Entwicklungssprunge“ und der enormen
Koharenz
der Evolution gesehen, sodass dieser ?Entscheidungsfreiheit der Materie“ eine maßgebliche Rolle als ?Treiber der Evolution“ gesehen wird. ?Nur geistige Eigenschaften konnen das Fundament der Wirklichkeit zusammenhalten und ihre stete evolutionare Entwicklung plausibel machen.“
[42]
schreibt Spat. Vielfach wird diese Freiheit in indeterminierten Prozessen auf der Quantenebene verortet,
[40]
wie sie etwa mit der
Hypothese
des
Quantendarwinismus
der Forschungsgruppe um
Wojciech Zurek
untersucht wird.
[46]
Nach Spat bietet der Selektionsmechanismus keine Erklarung fur die Entstehung geistiger Phanomene, weil etwa die Orientierung in der Umwelt
keine
Wahrnehmung voraussetzt (wie die
Robotik
beweist), sondern auch unbewusst und rein ?regelbasiert und physikalisch“ denkbar ist. Wo lage demnach der Uberlebensvorteil, den Vorgang der Orientierung zu
erleben
oder der Fahigkeit, eine vollig sinn- und wirkungslose (beim Menschen sogar eine bisweilen kontraproduktive, weil das Uberleben der Art gefahrdende) Innenwelt als ?Begleitmusik des Daseins“ zu ermoglichen? Hinzu kommt der enorme Energiebedarf geistiger Prozesse: Dass Nutzlichkeit und
Energieeffizienz
in aller Regel eine typische Folge evolutionarer Anpassungen sind, widerspricht der Entstehung solch ?nutzloser Zusatzfunktionen“.
[42]
Rekonstruktion des Evolutionsablaufs
Dieser Artikel oder nachfolgende Abschnitt ist nicht hinreichend mit
Belegen
(beispielsweise
Einzelnachweisen
) ausgestattet. Angaben ohne ausreichenden Beleg konnten demnachst entfernt werden. Bitte hilf Wikipedia, indem du die Angaben recherchierst und
gute Belege einfugst.
Die rekonstruierbaren Ablaufe der irdischen Evolution ? die Richtungen, die sie einschlug und die zeitlichen Einordnungen ? sind auf Basis des
Fossilberichts
sowie der Analyse rezenter Muster und Prozesse rekonstruierbar. Die ehemals abgelaufenen Prozesse bezuglich Richtung, zeitlicher Einordnung und Evolutionsgeschwindigkeit sind umso sicherer rekonstruierbar, je mehr unabhangige Indizien beitragen konnen, das historische Geschehen zu erhellen. Grundsatzlich wird vom
Aktualismus
fur den Ablauf geologischer und biologischer Prozesse der Vergangenheit ausgegangen, das heißt von der Annahme, dass die biologischen, okologischen und geologischen Prozesse in der Vergangenheit nach den gleichen oder ahnlichen Prinzipien abgelaufen sind, wie sie heute beobachtet und gemessen werden konnen. Hierzu konnen auch Experimente durchgefuhrt werden, die bis zu einem gewissen Grad auf Prozesse in der Vergangenheit projiziert werden konnen.
Bei Formen, die keine oder fast keine Fossilien hinterlassen haben, wozu fast alle
Prokaryoten
sowie die Mehrzahl der
eukaryotischen
Einzeller
gehoren, daneben auch alle skelettfreien sonstigen Organismen, wie
Wurmer
,
Quallen
,
Nacktschnecken
usw., konnen meist nur Vergleiche aus der rezenten (heutigen)
Fauna
oder
Flora
angestellt werden bei gleichzeitiger kritischer Plausibilitatsprufung der daraus abgeleiteten Schlussfolgerungen. Hier werden die Rekonstruktionen der zuruckliegenden Ablaufe vor allem auf Basis der molekularen Ahnlichkeit nach dem Analyseverfahren der phylogenetischen Verwandtschaft vorgenommen. Das Prinzip der
molekularen Uhr
kann helfen, die Abzweigungspunkte der verschiedenen Verwandtschaftslinien (
Kladen
) ungefahr zu datieren. Auch Prozesse der
Koevolution
, beispielsweise die ehemalige Aufnahme von Bakterien in Archaeen-Zellen, die dort (vor vielleicht zwei Milliarden Jahren, Abschatzung schwierig) zu den
Mitochondrien
und zu den
Chloroplasten
als
Endosymbionten
einer
eukaryotischen
Zelle geworden sind, konnen praktisch nur aus rezenten Daten uber
molekulargenetische
Ahnlichkeiten sowie Ahnlichkeiten im
Stoffwechsel
und in der Struktur der
Endosymbionten
erschlossen werden.
Am haufigsten sind skeletttragende Meeresformen als Fossilien zu finden, deutlich seltener Arten aus dem Festland von
Binnengewassern
(Flussen, Seen). Sehr wenige Fossilien hat man ublicherweise aus Gebirgsregionen, aus
Moor
- und Quellgewassern sowie auch generell aus ehemaligen Trockenzonen der Erde, da eine Einbettung und ein Erhalt an solchen Stellen generell eher unwahrscheinlich ist.
Fossile Uberlieferung
 Fossil eines
Archaeopteryx
.
Fossil eines
Archaeopteryx
.
Hinweise auf den zeitlichen Rahmen der Evolutionsablaufe geben die
Fossilien
, die
morphologisch
untersucht werden konnen, aus denen aber vielfach auch biologisch-okologische Eigenschaften, wie die besiedelten
Lebensraume
,
Bewegungsweisen
oder manchmal sogar das
Sozialverhalten
(z. B. wenn sie in
Rudeln
bzw.
Schwarmen
auftreten) ablesbar sein kann. Entsprechend der vertikalen Aufeinanderfolge fossilfuhrender
Gesteinsschichten
lassen sich Gemeinschaften fossiler Lebewesen (gemaß dem Prinzip der
Stratigraphie
) in eine zeitliche Reihenfolge bringen. Wahrend dies zunachst nur Informationen uber das ?relative Alter“ erbringt (welche Fossilien waren fruher, welche spater?), lasst sich mit Hilfe geeigneter
radiometrischer Methoden
in den Gesteinen und/oder den darin enthaltenen Fossilien eine Absolutdatierung vornehmen (
Geochronologie
).
Das Alter der Fossilien gibt Auskunft daruber, wann im Verlauf der
Stammesgeschichte
einzelner Gruppen sowie der Lebewesen insgesamt bestimmte Innovationen und
Aufspaltungsereignisse (adaptive Radiationen)
auftraten. Durch datierbare Fossilfunde ist zum Beispiel bekannt, dass (bis auf wenige Ausnahmen wie z. B.
Cloudina
)
[47]
alle Gruppen
skeletttragender
Tiere in einem engen Zeitfenster im fruhen bis mittleren
Kambrium
vor etwa 540 bis 500 Millionen Jahren erstmals auftraten.
[48]
Wie weit dies durch biologische Innovationen hervorgerufen wurde oder wie weit sich die Umweltbedingungen derart anderten, dass nunmehr Skelettbildungen und -ablagerungen chemisch-physikalisch moglich wurden, ist weiterhin eine nicht abschließend geklarte Frage.
Fossile Ubergangsformen (
Mosaikformen
und
Missing Links
) sind ein von Untersuchungen an rezenten Lebewesen unabhangiger Beweis fur die Verwandtschaft zwischen systematischen Großgruppen. Beruhmte Beispiele dafur sind ?
gefiederte Dinosaurier
“,
Archaeopteryx
und die Vogel der
Jehol-Gruppe
als Ubergangsformen von den nicht fliegenden
Amnioten
(umgangssprachlich ?Reptilien“ bzw. ? klassische ? ?Dinosaurier“, korrekt ?Nicht-Vogel-Dinosaurier“ genannt, beides
paraphyletische
Einheiten) und den modernen Vogeln
[49]
sowie
Panderichthys
,
Tiktaalik
und
Ichthyostega
als Ubergangsformen zwischen
Knochenfischen
und
Landwirbeltieren
.
[50]
Im gunstigsten Fall ist der Ubergang zwischen der ursprunglichen und der daraus abgeleiteten Gruppe durch eine Abfolge von Fossilfunden belegt, die mit abnehmendem Alter der neuen Gruppe morphologisch immer ahnlicher werden.
Evolutionare Trends sind in vielen Fallen innerhalb systematischer Gruppen gut dokumentiert, so z. B. bei den
pferdeartigen
Saugetieren: Aus vielzehigen, fuchsgroßen, laubfressenden Formen im
Alttertiar
sind uber mehrere Zwischenstufen die heutigen
Pferde
hervorgegangen.
Schließlich dokumentiert die fossile Uberlieferung Ab- und Zunahmen in der
Diversitat
systematischer Gruppen.
Faunenschnitte
sind
Massenaussterben
, bei denen in geologisch gesehen kurzen Zeitraumen die Zahl fossil uberlieferter
Taxa
stark reduziert wurde und manche Großgruppen vollig verschwanden oder in ihrer Vielfalt stark abnahmen. Bekanntester, wenngleich nicht großter Faunenschnitt ist das weitgehende Aussterben der ?
Dinosaurier
“ (mit Ausnahme der Gruppe der Vogel) und weiterer Großgruppen am Ende der
Kreidezeit
. Die freigewordenen okologischen Nischen konnten in der Folgezeit im Rahmen einer
Radiation
durch die Saugetiere und die
modernen Vogel
eingenommen werden.
Ein Uberblick uber das Auftreten der Tier- und Pflanzenstamme wird unter
Evolutionsgeschichte
prasentiert.
Phylogenetische Systematik
Der Merkmalsvergleich der Organismen im Rahmen der biologischen
Systematik
zeigte, dass die
Merkmale
nicht in beliebigen Kombinationen auftreten, sondern in einem System abgestufter Ahnlichkeiten. Dabei lassen sich Merkmalsgruppen gegeneinander abgrenzen, anhand derer die rezenten Organismen in Gruppen (
Taxa
, Einzahl
Taxon
), zusammengefasst (klassifiziert) und hierarchisch geordnet werden konnen.
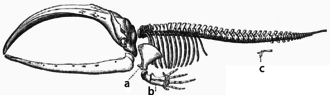 Skelett eines
Bartenwals
. Das
c
kennzeichnet die Rudimente der Hinterbeine; ein Merkmal, das die Abstammung von landlebenden Saugern belegt.
Skelett eines
Bartenwals
. Das
c
kennzeichnet die Rudimente der Hinterbeine; ein Merkmal, das die Abstammung von landlebenden Saugern belegt.
Dass dies moglich ist, wurde von Darwin als starkes Indiz fur eine gemeinsame Abstammung aller Lebewesen gewertet. Damit diese Gruppierungen (Systematisierungen) die tatsachlichen Verwandtschaftsverhaltnisse wiedergeben (
phylogenetische Systematik
), wurde als zentrales Kriterium die
Homologie
von Merkmalen eingefuhrt, das heißt, dass die Ahnlichkeit nur dann einen Aussagewert hat, wenn die entsprechenden Merkmale oder Organe auf die gleichen ursprunglichen Merkmale zuruckzufuhren sind. Die teilweise durch ihre
Sukkulenz
ahnlich aussehenden
Kakteen
Amerikas und
Wolfsmilchgewachse
Afrikas zeigen analoge Bildungen. Ihre oft ahnliche Erscheinungsform ist kein Hinweis auf eine engere Verwandtschaft und Abstammung von einem gemeinsamen Vorfahren.
Ein Sonderfall homologer Merkmale sind morphologische Merkmale oder sogar Verhaltensweisen, die fur ihre heutigen Trager keinen erkennbaren Zweck mehr erfullen (
Rudimente
), beispielsweise die Reste des Hinterbeinskeletts bei
Riesenschlangen
und
Walen
. In beiden Fallen weisen diese Rudimente auf die Abstammung von vierbeinigen Tieren (
Echsen
bzw.
Paarhufern
) hin.
Reste fruherer funktionierender Gene, die heute offensichtlich funktionslos sind, findet man im Genom beispielsweise bei manchen
Pseudogenen
.
Vergleichende Biogeographie
Die Verbreitung von Taxa liefert in vielen Fallen Hinweise fur evolutionare Entwicklungen. Viele Taxa haben geografische Verbreitungen, die allein durch die heutigen lokalen okologischen Verhaltnisse oder Anpassungen nicht zu erklaren sind. Dies gilt insbesondere fur
Endemiten
. Ein bekanntes Beispiel fur
Reliktendemiten
sind die
Lemuren
, ein Taxon innerhalb der
Feuchtnasenaffen
. Lemuren waren im Tertiar in Nordamerika und Eurasien weit verbreitet, sind heute aber auf
Madagaskar
endemisch. Sie wurden von den spater evolvierten und offensichtlich konkurrenzstarkeren
Trockennasenaffen
(Haplorhini) uberall verdrangt und konnten sich nur auf Madagaskar halten, das aufgrund einer zwischenzeitlich erfolgten geographischen Isolation von Trockennasenaffen nicht besiedelt werden konnte.
 Verschiedene Darwinfinken
Verschiedene Darwinfinken
Ebenso wichtig fur die Evolutionsbiologie sind
Entstehungsendemiten
. Beruhmtestes Beispiel sind hier die
Darwinfinken
(Familie Emberizidae) auf den Galapagosinseln, 965 km vor der Kuste
Ecuadors
. Auf der Inselgruppe vulkanischen Ursprungs, deren alteste Insel vor funf bis zehn Millionen Jahren entstand, entwickelten sich aus einer vom Festland verdrifteten Vorgangerart durch
adaptive Radiation
14 verschiedene Arten in drei Gattungen. Dabei anderten sich vor allem die Schnabelform, das Federkleid und die Große der Vogel. Charles Darwin sammelte die unterschiedlichen Arten 1836 auf seiner Weltreise.
Vergleichende Embryonalentwicklung
Karl Ernst von Baer
hat als erster erkannt, dass fruhe Entwicklungsstadien verwandter Organismen einander ahnlicher sind als die ausgewachsenen Individuen.
Ernst Haeckel
formulierte auf Basis dieser Erkenntnisse, dass die
Ontogenese
die Evolution (
Phylogenese
) eines Lebewesens rekapituliert (
Biogenetische Grundregel
). Diese Regel wird heute jedoch als zu starke Vereinfachung betrachtet. Wiederholt werden nicht die Adultstadien, sondern die fruhen
Embryonalstadien
der jeweiligen Vorformen einer Art. Das kann dadurch erklart werden, dass evolutionare Neuentwicklungen immer auf bestehende Arten und damit auf bestehende Organsysteme aufbauen. Daher schließt die Entwicklung (die Ontogenese) des Individuums einer Art auch die vorher jeweils bereits vorhandenen Schritte mit ein.
Beispiele einer solchen Wiederholung sind das Auftreten eines
Kiemendarms
und von Kiemenspalten bei allen
Wirbeltierembryonen
. Dies belegt, dass alle heutigen Wirbeltiere von Fischen abstammen.
Bartenwalembryonen
bilden Zahnanlagen, wahrend die
adulten
Individuen keine Zahne mehr haben, was darauf hinweist, dass sich die Bartenwale aus
Zahnwalen
entwickelt haben.
Offentliche Wahrnehmung
Dass eine Evolution im Sinne eines evolutiven Artenwandels und im Zusammenhang mit regelmaßigen Aussterbeereignissen auftrat, wurde seit jeher von einzelnen Personen oder von einem mehr oder weniger großen Teil der Bevolkerung in Abrede gestellt, wobei es lediglich in der fruhen Phase auch um wissenschaftliche Argumente ging. Uberwiegend geht es um grundsatzliche Haltungen auf Basis eines vollig anders konzipierten Weltbildes, das haufig auch den Planeten Erde selbst als nur einige tausend Jahre alt betrachtete. Manchmal wurden Einzelaspekte, wie ein ein- oder mehrmaliges
Massenaussterben
, das dann als mit den religios inspirierten
Sintflutberichten
in Einklang gebracht wird, akzeptiert, nicht aber andere Komponenten und Konsequenzen.
Wo das Prinzip der Evolution anerkannt wurde, wurden Teilaspekte davon in den letzten etwa 150 Jahren aber auch losgelost von
ethischen
Prinzipien als Mandat und Maxime eines politischen Handelns in der menschlichen Gesellschaft interpretiert und gefordert. Besonders im Fokus stand das Konzept eines universellen und auch die menschlichen Gesellschaften und ?
Rassen
“ umfassenden ?Kampfes ums Uberleben“, der aus dem Selektionsgedanken abgeleitet wurde und zur Haltung im
Sozialdarwinismus
fuhrte. Eine besondere Bedeutung erlangte in diesem Zusammenhang auch die Idee der
Eugenik
, die ab dem letzten Viertel des 19. Jahrhunderts im angelsachsischen Bereich popular wurde und dort beispielsweise die Einwanderungspolitik mit bestimmte. Nach Ubernahme der zugrunde liegenden Theorien und Ansichten auch in andere Staaten und in deren politische Fuhrungen kam es vor allem in Deutschland zu den radikalsten und folgenschwersten Auswirkungen zur
Zeit des Nationalsozialismus
(1933?1945). Aufgrund des damit verbundenen Massenmordes wurde Evolution und die damit zusammenhangende Theorie langere Zeit speziell im deutschen Sprachraum wissenschaftlich uberwiegend ausgeblendet und wenig bearbeitet. Wesentliche der modernen Forschungsentwicklungen fanden uber Jahrzehnte nur noch im angelsachsischen Bereich statt.
Eine grundsatzliche Gegnerschaft des Prinzips einer Evolution irdischen Lebens, speziell soweit auch die Spezies bzw. Gattung
Mensch
selber samt ihren Eigenschaften, wie Bewusstsein und geistiger Kreativitat, in die Betrachtung einbezogen werden, wurde langere Zeit und wird erneut seit einigen Jahrzehnten verstarkt von wissenschaftsskeptischen Bevolkerungsgruppen vertreten. Sie halten eine allein den Naturgesetzen unterworfene Entwicklung zu einem Wesen wie dem Menschen fur entweder grundsatzlich unvereinbar mit ihrem Weltbild oder fur so unwahrscheinlich, dass sie eine ubergeordnete lenkende Instanz, einen
Gott
, postulieren, der entweder die Arten erschaffen hat oder mindestens den Ablaufprozess eingeleitet oder gelenkt hat. Diese fundamentale Kontroverse ist in Europa vor allem durch die Auswirkungen um den Streit uber die Vermittlung der Evolutionstheorie im schulischen Biologieunterricht mancher
US-Bundesstaaten
bekannt geworden. Die entsprechenden Vertreter argumentieren haufig mit dem exakten Wortlaut der
Schopfung
sgeschichte, wie sie im
alten Testament
formuliert ist und folgen ihm wortgenau. Diese Skepsis bezuglich der Ergebnisse der Evolutionstheorie wird als
Kreationismus
bezeichnet. Eine Spezialform des Kreationismus ist die US-amerikanische
Intelligent-Design
-Idee.
Die
katholische Kirche
außerte sich wiederholt zur Evolutionstheorie, so Papst
Pius XII.
in seiner
Enzyklika
Humani generis
, Papst
Johannes Paul II.
auf der Vollversammlung der
Papstlichen Akademie der Wissenschaften
und Papst
Benedikt XVI.
in seiner Predigt zur Amtseinfuhrung. Die Evolutionstheorie wird heute vom
Vatikan
als ?vereinbar mit dem christlichen Glauben“ bezeichnet,
[51]
siehe auch
Theistische Evolution
. Der Wiener
Erzbischof
und
Kardinal
Christoph Schonborn
sprach sich mit dem Satz ?Evolution kann wahr sein“ fur eine ?große Koalition“ von Biologie und Theologie aus. Das Bibelverstandnis des Kreationismus sei nicht jenes der katholischen Kirche.
[52]
In der
fruhislamischen
wissenschaftlichen Blutezeit sind, teilweise in Anlehnung an griechische Vorbilder, darunter namentlich
Aristoteles
, Tierbeschreibungen und auch Gedankengange formuliert worden, die bereits an die evolutionsbiologischen Diskussionen des 19. Jahrhunderts erinnern, darunter ?
lamarckistische
“ Umwelteinflusse auf die Evolution, aber auch uber einen ?Kampf ums Dasein“ wurde spekuliert, was an das Selektionsprinzip Charles Darwins erinnert. Im 14. Jahrhundert sticht vor allem der islamische Gelehrte
Ibn Chald?n
hervor, der schrieb, dass sich Arten von Lebewesen durch naturliche Prozesse in neue Arten verwandeln. Weitere
islamische Gelehrte
wurden diskutiert, ihre Aussagen sind jedoch zu vage.
[53]
Im derzeitigen, durch stark divergierende Stromungen gekennzeichneten Islam gibt es keine einheitliche Meinung daruber, ob und in welchem Maße Evolution mit der Religion vereinbar sei; eine evolutionsskeptische Haltung ist relativ weit verbreitet:
[54]
Sowohl in der
Turkei
[55]
als auch in
Indien
[56]
wurde die Evolutionstheorie aus den Schullehrplanen entfernt. In etlichen islamisch gepragten Landern ist eine große Mehrheit der Menschen auch davon uberzeugt, dass sich die heutige Spezies
Homo sapiens
nicht evolutionar entwickelt hat, sondern statisch ist und von Gott (
Allah
) erschaffen wurde.
[57]
Die
literaturtheoretische
Schule des
Russischen Formalismus
griff um 1915 das Prinzip der biologischen Evolution auf, ubertrug es auf die
Literaturgeschichte
und entwickelte das Konzept der
Literarischen Evolution
.
Literatur
- Richard Dawkins:
The Selfish Gene.
Reissued in new covers. Oxford University Press, Oxford 1999,
ISBN 0-19-286092-5
(dt.:
Das egoistische Gen
).
- David J. Depew, Bruce H. Weber:
Darwinism Evolving. Systems Dynamics and the Genealogy of Natural Selection.
First MIT Press, Cambridge MA, 2. Aufl. 1997.
- Douglas J. Futuyma
, Mark Kirkpatrick:
Evolution.
4. Auflage. Sinauer 2017,
ISBN 978-1-60535-605-1
.
- Ulrich Kutschera
:
Evolutionsbiologie.
3. Auflage. Verlag Eugen Ulmer, Stuttgart 2008,
ISBN 978-3-8252-8318-6
.
- Ernst Mayr
:
Artbegriff und Evolution.
Parey-Verlag, Hamburg 1967.
- Ernst Mayr:
Das ist Evolution.
Goldmann, Munchen 2005,
ISBN 3-442-15349-2
.
- Volker Storch
,
Ulrich Welsch
,
Michael Wink
:
Evolutionsbiologie.
3. Auflage. Springer Spektrum Verlag, Heidelberg 2013,
ISBN 978-3-642-32835-0
.
Weblinks
- Literatur von und uber Evolution
im Katalog der
Deutschen Nationalbibliothek
- AG Evolutionsbiologie
Internetbibliothek mit zahlreichen Texten und ausgewahlter Fachliteratur zur Evolution sowie Literaturempfehlungen
- Evolution ? Zufall und Zwangslaufigkeit der Schopfung
Audiomitschnitte der offentlichen Ringvorlesung an der Universitat Gottingen im Wintersemester 2007/08
- Henry Gee, Rory Howlett and Philip Campbell ? 15 evolutionary gems
(PDF; 357 kB) Artikel uber 15 Falle der aktuellen Forschung, die die Darwin’sche Evolutionstheorie belegen. nature, 2009.
doi:10.1038/nature07740
.
- www.talkorigins.org
Englischsprachige Sammlung der haufigsten Missverstandnisse bezuglich der Evolutionstheorie und wie Wissenschaftler darauf antworten
- Geheimnisse der Evolution - Leben im Wasser.
Dokumentation.
ZDFinfo
,
abgerufen am 18. Januar 2022
(Synchronfassung ZDF 2019. Ein Film von Pierre-Francois Gaudry. Unter Mitwirkung von Guillaume Lecointre, Sarah Gabbot, Gavin Young, Bradley Smith, Andrew Gillis, John Long und Patrick Tschupp).
- Geheimnisse der Evolution - Leben an Land.
Dokumentation.
ZDFinfo
,
abgerufen am 19. Januar 2022
(Synchronfassung ZDF 2019. Ein Film von Pierre-Francois Gaudry. Unter Mitwirkung von Vera Weisbecker, Mihaela Parlicev, Guillaume Lecointre, Bradley Smith, Heather Smith, Michel Laurin, William Parker, Antoine Balzeau, Sergio Almecija, Henrik Kaessmann und Josh Griffiths).
Einzelnachweise
- ↑
F. D. Ciccarelli, T. Doerks, C. von Mering, C. J. Creevey, B. Snel, P. Bork:
Toward automatic reconstruction of a highly resolved tree of life.
In:
Science
.
Band 311, Nr. 5765, 2006, S. 1283?1287,
PMID 16513982
.
- ↑
A. Stoltzfus:
Mutationism and the dual causation of evolutionary change.
In:
Evol. Dev.
Band 8, Nr. 3, 2006, S. 304?317.
- ↑
zum Beispiel Ulrich Kutschera und
Karl J. Niklas
:
The modern theory of biological evolution: an expanded synthesis.
In:
Naturwissenschaften.
Band 91, Nr. 6, 2004, S. 255?276,
doi:10.1007/s00114-004-0515-y
.
- ↑
Helen Pearson:
Genetics: what is a gene?
In:
Nature.
Band 441, Nr. 7092, 2006, S. 398?401,
doi:10.1038/441398a
.
- ↑
Richard Mayeux:
Mapping the new frontier: complex genetic disorders.
In:
The Journal of Clinical Investigation.
Band 115, Nr. 6, 2005, S. 1404?1407,
doi:10.1172/JCI2542
.
- ↑
James A. Draghi, Paul E. Turner:
DNA secretion and gene-level selection in bacteria.
In:
Microbiology.
Band 152, Nr. 9, 2006, S. 2683?2688,
doi:10.1099/mic.0.29013-0
.
- ↑
James Mallet:
Hybrid speciation.
In:
Nature
.
Band 446, Nr. 7133, 2007, S. 279?283,
doi:10.1038/nature05706
.
- ↑
R. K. Butlin, T. Tregenza:
Levels of genetic polymorphism: marker loci versus quantitative traits.
In:
Philosophical Transactions of the Royal Society
, B, Biological Sciences.
Band 353, Nr. 1366, 1998, S. 187?198,
doi:10.1098/rstb.1998.0201
,
PMC 1692210
(freier Volltext).
- ↑
A. Wetterbom, M. Sevov, L. Cavelier, T. Bergstrom:
Comparative genomic analysis of human and chimpanzee indicates a key role for indels in primate evolution.
In:
Journal of Molecular Evolution.
Band 63, Nr. 5, 2006, S. 682?690,
doi:10.1007/s00239-006-0045-7
.
- ↑
Roy J. Britten
:
Divergence between samples of chimpanzee and human DNA sequences is 5 %, counting indels.
In:
PNAS
.
Band 99, Nr. 21, 2002,S. 13633?13635,
doi:10.1073/pnas.172510699
.
- ↑
William Amos, John Harwood:
Factors affecting levels of genetic diversity in natural populations.
In:
Philosophical Transactions of the Royal Society of London, B, Biological Sciences.
Band 353, Nr. 1366, 1998, S. 177?186,
doi:10.1098/rstb.1998.0200
,
PMC 1692205
(freier Volltext).
- ↑
M. Andersson, L. Simmons:
Sexual selection and mate choice.
In:
Trends in Ecology and Evolution.
Band 21, Nr. 6, 2006, S. 296?302.
- ↑
zum Beispiel J. Hunt, R. Brooks, M. Jennions, M. Smith, C. Bentsen, L. Bussiere:
High-quality male field crickets invest heavily in sexual display but die young.
In:
Nature.
Band 432, Nr. 7020, 2004, S. 1024?1027.
- ↑
J. R. Krebs, N. B. Davies:
Einfuhrung in die Verhaltensokologie
Blackwell, 1996, S. 343 ff.
- ↑
Richard Dawkins:
The Selfish Gene
.
1976.
- ↑
J. R. Krebs, N. B. Davies:
Einfuhrung in die Verhaltensokologie.
Blackwell, 1996, S. 387 ff.
- ↑
R. Lande:
Fisherian and Wrightian theories of speciation.
In:
Genome.
Band 31, Nr. 1, 1989, S. 221?227.
- ↑
M. Whitlock:
Fixation probability and time in subdivided populations.
In:
Genetics.
Band 164, Nr. 2, 2003, S. 767?779.
- ↑
S. Otto, M. Whitlock:
The probability of fixation in populations of changing size.
In:
Genetics.
Band 146, Nr. 2, 1997, S. 723?733.
- ↑
Andreas Werner, Monica J. Piatek, John S. Mattick:
Transpositional shuffling and quality control in male germ cells to enhance evolution of complex organisms
. In:
Annals of the New York Academy of Sciences
. 1341. Jahrgang,
Nr.
1
, April 2015,
S.
156?163
,
doi
:
10.1111/nyas.12608
,
PMID 25557795
,
PMC 4390386
(freier Volltext),
bibcode
:
2015NYASA1341..156W
.
- ↑
Christoph Adami:
What is complexity?
In:
BioEssays
. 24. Jahrgang,
Nr.
12
, 22. November 2002,
S.
1085?1094
,
doi
:
10.1002/bies.10192
,
PMID 12447974
.
- ↑
M. Mitchell Waldrop; N. Dewar (Zeichnungen):
Language: Disputed definitions. §Complexity
. In: Heidi Ledford (Hrsg.):
Nature
. 455. Jahrgang,
Nr.
7216
, 23. Oktober 2008,
S.
1023?1028
,
doi
:
10.1038/4551023a
,
PMID 18948925
(englisch).
- ↑
a
b
c
d
Giuseppe Longo, Mael Montevil:
Randomness Increases Order in Biological Evolution
. Hrsg.: Michael J. Dinneen, Bakhadyr Khoussainov, Andre Nies (=
Computation, Physics and Beyond. Lecture Notes in Computer Science (LNTCS)
.
Band
7160
). Springer Berlin Heidelberg, 2012,
ISBN 978-3-642-27653-8
,
S.
289?308
,
doi
:
10.1007/978-3-642-27654-5_22
(englisch,
Online
).
SemanticScholar
,
Randomness Increases Order in Biological Evolution
. Siehe insbes.
Fig. 2
Evolution der Komplexitat nach
Stephen J. Gould
. Diese Darstellung erfolgt auf der Grundlage palaontologischer Beobachtungen.
- ↑
Pascal Sirand-Pugnet, Carole Lartigue, Marc Marenda, Daniel Jacob, Aurelien Barre, Valerie Barbe, Chantal Schenowitz, Sophie Mangenot, Arnaud Couloux, Beatrice Segurens, Antoine de Daruvar, Alain Blanchard, Christine Citti:
Being Pathogenic, Plastic, and Sexual while Living with a Nearly Minimal Bacterial Genome
. In:
PLOS Genet.
3. Jahrgang,
Nr.
5
, 18. Mai 2007,
S.
e75
,
doi
:
10.1371/journal.pgen.0030075
,
PMID 17511520
,
PMC 1868952
(freier Volltext).
- ↑
Heather Maughan, Joanna Masel, C. William Birky Jr., Wayne L. Nicholson:
The roles of mutation accumulation and selection in loss of sporulation in experimental populations of Bacillus subtilis
. In:
Genetics
. 177. Jahrgang,
Nr.
2
, 1. Oktober 2007,
S.
937?948
,
doi
:
10.1534/genetics.107.075663
,
PMID 17720926
,
PMC 2034656
(freier Volltext).
- ↑
Michael Ruse
:
Monad to man: the Concept of Progress in Evolutionary Biology
. Harvard University Press, 1996,
ISBN 978-0-674-03248-4
,
S.
526?529
(
Webarchive
).
- ↑
a
b
Sean B. Carroll:
Chance and necessity: the evolution of morphological complexity and diversity
. In:
Nature
. 409. Jahrgang,
Nr.
6823
, 22. Februar 2001,
S.
1102?1109
,
doi
:
10.1038/35059227
,
PMID 11234024
,
bibcode
:
2001Natur.409.1102C
.
- ↑
a
b
c
Stephen Jay Gould
:
The Evolution Of Life On Earths
. In: Scientific American, Oktober 1994 (via brembs.net). Siehe insbes. Fig. 2: Progress Does Not Rule. Dazu:
- ↑
a
b
c
Stephen Jay Gould
:
Full House: The Spread of Excellence from Plato to Darwin
. Harmony Books, USA 1996, 244 Seiten,
ISBN 978-0-517-70394-6
. Three Rivers Pr. 1997, S. 171. Dazu:
- ↑
Aharon Oren:
Prokaryote diversity and taxonomy: current status and future challenges
. In:
Philos. Trans. R. Soc. Lond. B Biol. Sci.
359. Jahrgang,
Nr.
1444
, 29. April 2004,
S.
623?638
,
doi
:
10.1098/rstb.2003.1458
,
PMID 15253349
,
PMC 1693353
(freier Volltext).
- ↑
Robert Mccredie May, R. J. H. Beverton:
How Many Species?
In:
Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences
. 330. Jahrgang,
Nr.
1257
, 29. November 1990,
S.
293?304
,
doi
:
10.1098/rstb.1990.0200
.
- ↑
Patrick D. Schloss, Jo Handelsman:
Status of the microbial census
. In:
Microbiol Mol Biol Rev
. 68. Jahrgang,
Nr.
4
, 1. Dezember 2004,
S.
686?91
,
doi
:
10.1128/MMBR.68.4.686-691.2004
,
PMID 15590780
,
PMC 539005
(freier Volltext).
- ↑
Noel A. Heim, Jonathan Louis Payne, Seth Finnegan, Matthew L. Knope, Michał Kowalewski, S. Kathleen Lyons, Daniel W. McShea, Philip M. Novack-Gottshall, Felisa A. Smith, Steve C. Wang:
Hierarchical complexity and the size limits of life
. In: Proc. R. Soc. B., Band 284, Nr. 1857; 21. Juni 2017, S. 20171039;
doi:10.1098/rspb.2017.1039
,
PMID 28637850
,
PMC 5489738
(freier Volltext),
ResearchGate
. Siehe insbes.
Fig. 1
: Aggregated distributions of organismal size. (a) The hypothetical unimodal right-skewed distribution of organismal sizes expected under the Gould model (Aggregierte Verteilungen der Organismengroße. (a) Die hypothetische unimodale rechtsschiefe Verteilung der Organismengroßen, die nach dem Gould-Modell erwartet wird).
- ↑
Jesse M. Mulder:
Die Grenzen des Reduktionismus: Denken, Leben und Wirklichkeit.
2000, unterstutzt durch die niederlandische Nationale Wissenschaftliche Stiftung NWO, VENI Grant (Nummer 275-20-068),
PDF
abgerufen am 25. Februar 2024, S. 2?5, 12.
- ↑
Thomas Diekwisch:
Biologische Beitrage zum Leib-Seele-Problem.
Inauguraldissertation, Philipps-Universitat Marburg, Bielefeld 2004,
PDF
, abgerufen am 25. Februar 2024, S. 259?260, 275, 290.
- ↑
Ralph Adolphs und
Antonio R. Damasio
:
Bewusstsein und Neurowissenschaft.
DOI:10.1515/9783110525601-010
, in: Burkhart Bromm, Jorn Henning Wolf (Hrsg.):
Von der Freiheit, Schmerz zu spuren.
Abhandlungen der Akademie der Wissenschaften in Hamburg, Band 7. de Gruyter, Berlin/Boston 2017,
ISBN 978-3-11-052351-5
, S. 77?78, 83, 90?93.
- ↑
Edwin Egeter:
Phanomenale Adaquatheit und Irreduzibilitat des Bewusstseins. Eine Revision des Qualia-Begriffs.
mentis/brill, Paderborn 2020,
ISBN 978-3-95743-194-3
.,
PFD
, abgerufen am 9. Februar 2024, S. 334?335, 339.
- ↑
Richard H. Beyler:
Physik.
In:
Philipp Sarasin
,
Marianne Sommer
(Hrsg.):
Evolution. Ein interdisziplinares Handbuch
. J. B. Metzler, Stuttgart/Weimar 2010,
ISBN 978-3-476-02274-5
, S. 293.
- ↑
Samuel R. Nuesch: Die Leib-Seele Debatte: Eine Ubersicht der wichtigsten Positionen. Arbeitspapiere aus der IKAO, Nr. 1, Universitat Bern, September 2008, [www.ikaoe.unibe.ch/publikationen/arbeitspapier_01.pdf PDF], abgerufen am 22. Februar 2024, S. 5, 16?18.
- ↑
a
b
c
d
Michael Hampe
:
Philosophie.
In:
Philipp Sarasin
,
Marianne Sommer
(Hrsg.):
Evolution. Ein interdisziplinares Handbuch
. J. B. Metzler, Stuttgart/Weimar 2010,
ISBN 978-3-476-02274-5
, S. 273?284.
- ↑
Manfred Stockler
:
Gehirn, Bewusstsein und Schmerz ? eine Skizze, wie alles zusammenpassen konnte.
DOI:10.1515/9783110525601-011
, in: Burkhart Bromm, Jorn Henning Wolf (Hrsg.):
Von der Freiheit, Schmerz zu spuren.
Abhandlungen der Akademie der Wissenschaften in Hamburg, Band 7. de Gruyter, Berlin/Boston 2017,
ISBN 978-3-11-052351-5
, S. 101?104.
- ↑
a
b
c
Patrick Spat
:
Panpsychismus: ein Losungsvorschlag zum Leib-Seele-Problem.
Dissertation, FreiDok der Universitat Freiburg, Freiburg 2010,
PDF
, abgerufen am 17. Juni 2023, S. 2?4, (Inspiration von Whitehead u. Teilhard), 4, 91?93, (Evolution), 81?86 (Emergenz), 92?93 (Evolutionarer Vorteil von Bewusstsein).
- ↑
Heinrich Pas:
Gibt es den freien Willen? Und: Was ist Realitat?
Beitrag auf Spektrum.de SciLogs vom 16. Januar 2018,
online
abgerufen am 29. Februar 2024.
- ↑
Robert Prentner:
Die Entstehung der Objekte. Uberlegungen zu einer exakten Wissenschaft von Bewusstsein.
Dissertation ETH Zurich, Nr. 24329, 2017,
PDF
, abgerufen am 29. Februar 2024, S. 51.
- ↑
Bernhard Rensch, u. a.:
Biophilosophie auf erkenntnistheoretischer Grundlage (Panpsychistischer Identismus)
, Stuttgart 1968.
- ↑
Florian Aigner
:
Warum wir nicht durch Wande gehen
. Brandstatter, Wien 2023,
ISBN 978-3-7106-0689-2
, S. 203?208.
- ↑
H. Hua, Z. Chen, X. Yuan, L. Zhang, S. Xiao:
Skeletogenesis and asexual reproduction in the earliest biomineralizing animal 'Cloudina'.
In:
Geology.
Band 33. Nr. 4, 2005, S. 277?280.
- ↑
?
kambrische Explosion
“: Review in S. Conway Morris:
The fossil record and the early evolution of the metazoa.
In:
Nature.
Band 361, Nr. 6409, 1993, S. 219?225.
- ↑
Review-Artikel zum Dinosaurier-Vogel-Ubergang:
M. A. Norell, X. Xu:
Feathered dinosaurs.
In:
Annual Review of Earth and Planetary Sciences.
Band 33, 2005, S. 277?299.
F. Zhang, Z. Zhou, G. Dyke:
Feathers and ?feather-like‘ integumentary structures in liaoning birds and dinosaurs.
In:
Geological Journal.
Band 41, Nr. 3?4, 2006, S. 395?404.
Z. Zhou:
The origin and early evolution of birds: Discoveries, disputes, and perspectives from fossil evidence.
In:
Naturwissenschaften.
Band 91, Nr. 10, 2004, S. 455?471.
Z. Zhou, F. Zhang:
Mesozoic birds of china ? A synoptic review.
In:
Frontiers of Biology in China.
Band 2, Nr. 1, 2007, S. 1?14.
- ↑
J. A. Long, G. C. Young, T. Holland, T. J. Senden, E. M. G. Fitzgerald:
An exceptional devonian fish from australia sheds light on tetrapod origins.
In:
Nature.
Band 444, Nr. 7116, 2006, S. 199?202.
- ↑
Chris Irvine:
The Vatican claims Darwin's theory of evolution is compatible with Christianity.
In:
telegraph.co.uk.
11. Februar 2009,
abgerufen am 30. Mai 2015
(englisch).
- ↑
Schonborn fur "Große Koalition" von Biologie und Theologie.
Die Presse, 5. Marz 2009,
abgerufen am 12. Oktober 2020
.
- ↑
Turgut Demirci:
Die Vereinbarkeit der wissenschaftlichen Evolutionstheorie mit dem Islam.
Masterarbeit, Universitat Wien 2016,
PDF
abgerufen am 3. November 2023, S. 93?112
(4.2. Die Evolutionsidee bei muslimischen Denkern vor Darwin)
, 104?106
(4.2.4. Ibn Khaldun und die Umwandlung der Arten)
.
- ↑
SWR 2009: Islam und Darwin ? ein Widerspruch ? ? abgerufen am 15. Februar 2016
(
Memento
vom 1. Marz 2016 im
Internet Archive
)
- ↑
Deutschlandradio:
Darwins Lehre wird vom Lehrplan gestrichen.
Abgerufen am 27. August 2023
.
- ↑
Humanistischer Pressedienst:
Wissenschaftsleugnung und Geschichtsklitterung im Dienste nationalistischer Politik.
Abgerufen am 27. August 2023
.
- ↑
PewResearchCenter:
The World’s Muslims: Religion, Politics and Society.
(
Memento
vom 30. Oktober 2014 im
Internet Archive
) 4/2013, S. 132.