Bovidae
perben nje familje biologjike thundrake, ripertypes
vertebrore
qe perfshin
bizonet
,
buajt afrikan
,
buallicat
,
antilopat
,
delet
,
dhite
,
buajt e myshqeve
, dhe
lopet
. Nje anetar i kesaj familje quhet bovid . Me 143 specie ekzistuese dhe 300 lloje te njohura te zhdukura, familja Bovidae perbehet nga tete nenfamiljet kryesore pervec Peleinae dhe Pantholopinae te diskutueshme. Familja evoluoi 20 milion vjet me pare, ne Miocenin e hershme .
Foshnjat tregojne nje ndryshim te madh ne madhesi dhe
gezof
nga te rriturit. Me perjashtim te disa formave shtepiake , te gjithe bovidet meshkulljt kane dy ose me shume
brire
, dhe ne shume specie, femrat gjithashtu posedojne brire. Madhesia dhe forma e brireve ndryshojne shume, por struktura themelore eshte gjithmone nje ose me shume cifte te zgjatjeve te thjeshta te eshtrave pa dege, shpesh qe kane nje forme spirale, te perdredhur ose te drejta, secila e mbuluar ne nje kellef te perhershem te keratines . Shumica e gjedheve kane 30 deri ne 32 dhembe.
Shumica e gjedheve jane lloje ditore . Aktiviteti shoqeror dhe ushqyerja zakonisht arrijne kulmin gjate
agimit
dhe
muzgut
. Bovidet zakonisht pushojne para agimit, gjate mesdites dhe pas erresires. Ato kane metoda te ndryshme te organizimit shoqeror dhe sjelljes shoqerore , te cilat klasifikohen ne sjellje te vetmuar. Bovidet perdorin forma te ndryshme te komunikimit vokal, nuhates dhe te prekjes. Shumica e specieve ushqehen dhe piqen ne menyre alternative gjate gjithe dites. Ndersa bovidet e vegjel ushqehen ne nje habitat te dendur dhe te mbyllur, speciet me te medha ushqehen me bimesi me fibra te larte ne kullota te hapura. Shumica e bovideve jane poligjen. Bovidet e pjekur bashkohen te pakten nje here ne vit dhe speciet me te vogla mund te bashkohen edhe dy here. Ne disa lloje, gjinjte e posalindur mbeten te fshehur per nje jave deri ne dy muaj, duke u kujdesur rregullisht nga nenat e tyre.
Shumellojshmeria me e madhe e bovideve ndodhen ne
Afrike
. Perqendrimi maksimal i specieve eshte ne savanat e
Afrikes Lindore
. Lloje te tjera te bovideve gjithashtu ndodhen ne
Evrope
,
Azi
dhe
Ameriken e Veriut
. Bovidae perfshin tre nga pese gjitaret shtepiak , perdorimi i te cilave eshte perhapur jashte kufijve te tyre origjinal, perkatesisht lopet, dele dhe dhi. Produktet e bulmetit si
qumeshti
,
gjalpi
dhe
djathi
prodhohen kryesisht nga lopet. Bovidet gjithashtu sigurojne
lekure
,
mish
dhe
gezof
.
Emri "Bovidae" u dha nga zoologu britanik
John Edward Grey
ne 1821.
[1]
Fjala "Bovidae" eshte nderthurja e parashteses
bov-
(me origjine nga
fjala latine
bos
,"dem" ) dhe prapashtesa
-idae
.
[2]
Familja Bovidae eshte vendosur ne rendin Artiodaktila (i cili perfshin
dythundraket
). Ai perfshin 143 specie te gjalla, qe perbejne gati 55% te thundrakeve , dhe 300 specie te njohura te zhdukura.
[3]
Studimet molekulare kane mbeshtetur ne menyre monofile ne familjen Bovidae (nje grup i organizmave perfshin nje specie stergjyshore dhe te gjithe pasardhesit e tyre).
[4]
[5]
Numri i nenfamiljeve tek Bovidae eshte i diskutueshem, me sugjerime te me shume se dhjete dhe sa me pak si dy nenfamilje.
[5]
Sidoqofte, provat molekulare, morfologjike dhe fosile tregojne ekzistencen e tete nenfamiljeve te dallueshme:
Aepycerotinae
(qe perbehet vetem nga
impala
),
Alcelaphinae
(antilopat afrikane), Antilopina (disa
antilopa
,
gazelat
dhe te aferm),
Bovinae
(
lopet
,
buallicat
,
bizonet
dhe antilopa te tjera),
Caprinae
(
dhite
,
delet
,
serout
dhe te afermit), Cephalophinae (duikeret),
Hippotraginae
(
adaksin
,
oriksin
dhe lloje te tjera) dhe
Reduncinae
(antilopat kob). Per me teper, tre nenfamilje te zhdukura jane te njohura: Hypsodontinae (mes Miocenit), Oiocerinae ( Turolianin ) dhe nenfamilja Tethytraginae, e cila permban
Tethytragus
(mes-Miocenit).
[6]
[7]
Ne vitin 1992, Alan W. Gentry i Muzeut te Historise Natyrore, Londer, ndau tete nenfamiljet kryesore te Bovidae ne dy kada kryesore mbi bazen e historise se tyre evolucionare: Boodontia, e cila perbente vetem Bovinae, dhe Aegodontia, e cila perbehej nga pjesa tjeter e nenfamiljeve. Boodontet kane dhembe disi primitive , te ngjashem me ato te lopeve, ndersa aegodonet kane dhembe me te perparuar si ato te dhive.
[8]
Kundershtimi ekziston per njohjen e Peleinae dhe Patholopinae, qe permban pergjithshme
Pelea
dhe
Pantholops
, sic nenfamilje. Ne vitin
2000
, biologu amerikan
George Schaller
dhe paleontologjisti
Elisabeth Vrba
sugjeroi perfshirjen e
Pelea
ne Reduncinae,
[9]
sepse kobi gri, eshte specie e vetme e
Pelea
, eshte shume i ndryshem nga kobet ne morfologji.
[10]
Pantholops
, ishte klasifikuar me pare ne Antilopina, por u vendos me vone ne nenfamiljen e vet, Pantholopinae. Sidoqofte, analiza molekulare dhe morfologjike mbeshtet perfshirjen e
Pantholops
ne Caprinae.
[11]
Me poshte eshte nje kladogrami bazuar nga Gatesy et al. (1997) dhe Gentry et al. (1997)
 Kafka e
Eotragus sansaniensis
, nje specie e gjinise se lashte
Eotragus
Kafka e
Eotragus sansaniensis
, nje specie e gjinise se lashte
Eotragus
Ne fillim te
Miocenit
, bovidet filluan divergjencen e tyre nga cervidet (
dreret
) dhe
girafidet
. Bovidet me te hershme, ishin ne Afrike dhe
Euroazi
ne pjesen e fundit te Miocenit te hershem (20 Milion ) eshte konstatuar,duke treguar se ishin kafshe te vogla, disi te ngjashme me gazelat moderne , dhe me siguri jetonin ne mjedise pyjore .
[12]
Eotragus
, bovidi me i hershem i njohur, peshonte 18 kg dhe ishte pothuajse i njejte me madhesine e
gazeles se Tompsonit
.
[13]
Ne fillim te historise se tyre evolucionare, bovidet u ndane ne dy kadra kryesore: Boodontia (me origjine Euroaziatike) dhe Aegodontia (me origjine Afrikane). Kjo ndarje e hershme midis Boodontia dhe Aegodontia i eshte atribuar ndarjes kontinentale midis ketyre masave tokesore. Kur me vone keto kontinente u ribashkuan, kjo pengese u hoq, dhe secili grup u zgjerua ne territorin e tjetrit.
[14]
Fiset Bovini dhe Tragelaphini ndryshuan ne Miocenin e hershem.
[15]
Bovidet dihet se kane arritur ne Ameriken ne Pleistocene duke kaluar uren tokesore te Beringit .
[13]
Nenfamilja e tanishme e Alcelaphinae u shfaq ne Pliocene . Lloji i zhdukur alcelafin
Paramularius
, i cili ishte i njejti me madhesine e kongonit , besohet te kete ardhur ne Pliocene, por u zhduk ne Pleistocenin e mesem .
[5]
Disa gjini te Hippotraginae jane te njohura qe nga Plioceni dhe Pleistoceni. Kjo nenfamilje duket se ka devijuar nga Alcelaphinae ne pjesen e fundit te Miocenit te hershem.
[15]
Bovinae besohet te kete devijuar nga pjesa tjeter e Bovidae ne fillim te Miocenit .
[16]
Boselafini u zhduk ne Afrike ne fillimin e poliocenit; fosilet e tyre te fundit u germuan brenda Langebaanweg (
Afrika e Jugut
) dhe Lothagam (
Kenia
).
[17]
Fosilet e hershme te antilopineve jane nga Mioceni i mesem, megjithese studimet tregojne ekzistencen e nenfamiljes nga Mioceni i hershem. Specifikimi ka ndodhur ne fisin Antilopini gjate Miocenit te mesem ose te vone, kryesisht ne Euroazi. Fisi Neotragini duket se jane shfaqur ne Afrike deri ne fund te Miocenit, dhe ishte bere e perhapur nga
plioceni
.
[15]
Ne fund te Miocenit, rreth 10 milion vite me pare, bovidet u diversifikuan me shpejtesi , duke cuar ne krijimin e 70 gjinive te reja.
[13]
Kjo shumllojshmeri i miocenit te vone ishte pjeserisht sepse shume bovidet u bene pershtatje per habitatet me te hapura, me barishte.
[12]
Aepycerotinae se pari u shfaq ne Miocenin e vone, dhe nuk eshte verejtur ndonje ndryshim i rendesishem ne madhesite e impales primitive dhe moderne.
[18]
Fosilet e ovibovineve, nje fis i Caprinae, ne Afrike datojne nga Mioceni i vone.
[15]
Fosilet e hershme te Hipotragineve datojne nga Mioceni i vone, dhe u germuan nga vendet si Lothagam dhe Lugina e Awash .
Fosilet
e para afrikane te Reduncinae datojne nga 6-7 miion vite me pare.
[15]
Peleinae ndoshta devijuan ne mes te Miocenit.
[19]
Reduncinae and Peleinae probably diverged in the mid-Miocene.
[5]
 Bovidet kane brire.
Bovidet kane brire.
Te gjithe bovidet kane forma te njejta - nje ose me shume brire (pergjithesisht te pranishem tek meshkujt) menjehere pas vesheve ovale ose te theksuar, nje qafe dhe gjymtyre te vecante, dhe nje bisht i ndryshem ne gjatesi dhe forme ne mesin e specieve.
[20]
Shumica e bovidet shfaqin dimorfizem seksual , me meshkuj zakonisht me te medhenj, si dhe me te rende se femrat.
Dimorfizmi seksual
eshte me i spikatur tek bovidet me madhesi mesatare dhe te medha. Te gjithe bovidet kane kater gishterinj ne secilen kembe - ato ecin me dy qendroret ( thundrat ), ndersa dy pjeset e jashtme ( rrobat e vese ) jane shume me te vogla dhe rralle prekin token.
[3]
Bovidet tregojne ndryshime te medha ne madhesi: trupi mund te peshoje me shume se 1000 kg dhe qendrojne 2-3 m i larte ne sup.
[21]
Buallica mund te jete me i rendi i bovideve, dhe peshon 1.200 kg , edhe pse ajo eshte me e shkurter se sa
Gauri
, duke qene 2 m i gjate. Antilope mbreterore , ne kontrast te mprehte, eshte vetem 25 cm i gjate dhe peshon me se shumti 3 kg .
[22]
Klipspringeri
, nje antilope e vogel, qendron 45-60 cm ne shpatullen dhe peshon vetem 10-20 kg .
[23]
Dallimet ndodhin ne ngjyrimin e gezofit, duke filluar nga nje zbehje te bardhe (si oriksi arab ) ne te zeze (si antilopa e zeze afrikane ).
[24]
[25]
Sidoqofte, zakonisht verehen vetem nuancat e ndermjetme, si e murrmja dhe kafeja e kuqerremte.
[26]
Ne disa lloje, femrat dhe te miturit shfaqin nje pallto me ngjyra te lehta, ndersa ato te meshkujve erresohen me moshen. Si tek antilopat afrikane , palltot mund te jete e shenuar me vija te spikatura ose te zbehta. Ne disa lloje te tilla si adaksi , ngjyra e palltos mund te ndryshoje sipas stines.
[27]
Gjendra aromatike dhe gjendra dhjamore jane shpesh te pranishme.
[20]
 Gemsboku ka shenja dukshem ne fytyren e tij, te cilat fshehin syrin, dhe ne kembet e saj. Keto mund te kene nje rol ne komunikim.
[28]
Gemsboku ka shenja dukshem ne fytyren e tij, te cilat fshehin syrin, dhe ne kembet e saj. Keto mund te kene nje rol ne komunikim.
[28]
Disa lloje, te tilla si
Gemsboku
,
antilopa Sablei
, dhe
gazela e Grantit
,mund te kamofloen. Shume specie, te tilla si gazelat, mund te behen qe te duken si bari i thate, dhe keshtu te perzihen ne sfond, duke u kundervene .
[29]
Skicat e shume bovideve jane te ndara me ngjyrosje te guximshme percarese, modele fort te kunderta qe ndihmojne per te vonuar njohjen nga grabitqaret. Sidoqofte, te gjithe Hippotraginae (perfshire gemsboku) kane trupa te zbehte dhe fytyra me shenja te dukshme.
[30]
Zoologu
Tim Caroe
pershkruan kete si te veshtire per tu shpjeguar, por duke pasur parasysh qe speciet jane ditore, ai sugjeron qe shenjat mund te funksionojne ne komunikim.
[31]
Ngjyrosja e forte e kundert e kembeve eshte e zakonshme vetem tek Bovidae, ku per shembull
Bos
,
Ovis
, bonteboku dhe gemsboku kane kembe te bardha. Perseri, komunikimi eshte funksioni i mundshem.
[28]
Me perjashtim te disa formave primitive, te gjitha specie e bovideve meshkujt kane brire, dhe ne shume specie, edhe femrat, posedojne brire. Madhesia dhe forma e brireve ndryshojne shume, por struktura themelore eshte nje cift zgjatimesh te thjeshta te eshtrave pa dege, shpesh qe kane nje forme spirale, te shtremberuar ose te rrjedhshme, secila e mbuluar ne nje kellef te perhershem te keratines. Megjithese briret ndodhen ne nje cift te vetem ne pothuajse te gjitha llojet e bovideve, ka perjashtime te tilla si antilope me kater brire
[32]
dhe delet e Jakobit .
[33]
[34]
Struktura unike e brireve eshte e vetmja vecori morfologjike e paqarte e gjedheve qe i dallon ato nga pekoranet e tjere .
[35]
[36]
Nje lidhje e larte ekziston midis morfologjise se bririt dhe sjelljes luftarake te individit.
[37]
Per shembull, briret e gjata jane te destinuara per mundje, ndersa briret e lakuar perdoren ne mbrotje. Meshkujt me brire te drejtuar nga brenda jane monogame dhe vetmuar, ndersa ata me brire te drejtuar nga jashte kane tendence te jene
poligjen
. Keto rezultate ishin te pavarura nga madhesia e trupit.
[38]
Zhvillimi i brireve tek meshkuj ka qene i lidhur me perzgjedhjen seksuale .
[39]
[40]
Briret jane thumba te vogla ne duikeret monogame dhe antilopat e tjera te vogla, ndersa ne poligjen, ato jane te medha dhe te formuara hollesisht (per shembull ne nje strukture spirale, si tek
Elandi gjigant
). Keshtu, ne nje fare mase, briret pershkruajne shkallen e konkurrences midis meshkujve ne nje specie.
[26]
Sidoqofte, prania e brireve tek femra ka te ngjare te jete per shkak te seleksionimit natyror .
[39]
[41]
Briret e femrave jane zakonisht me te vogla se ato te meshkujve, dhe ndonjehere jane te nje forme te ndryshme. Briret e bovideve femer besohet se kane evoluar per mbrojtje kunder grabitqareve ose per te shprehur territorin, pasi femrat jospitimprurese, te cilat jane ne gjendje te perdorin perzierien ne nature per mbrojtjen e grabitqareve, shpesh nuk kane brire. Femrat zoterojne brire vetem ne gjysmen e gjinive te bovideve, dhe femrat ne keto gjini jane me te renda se ato ne pjesen tjeter. Femrat perdorin briret kryesisht per thike.
[42]
 Skeleti i bizonit amerikan ( Muzeu i Osteologjise )
Skeleti i bizonit amerikan ( Muzeu i Osteologjise )
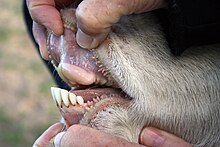 Jasteku i dhembeve i nje lope: Vini re mungesen e dhembeve te siperm dhe qeneve dhe projeksionin e jashtem te dhembeve te poshtem.
Jasteku i dhembeve i nje lope: Vini re mungesen e dhembeve te siperm dhe qeneve dhe projeksionin e jashtem te dhembeve te poshtem.
Tek bovidet, metapodialet e trete dhe te katert kombinohen ne
kocken
e topave . Ulna dhe kerci duke reduktuar, dhe te perziera me rrezen dhe kembet, respektivisht. Skapula te gjata jane te pranishme, ndersa klavikulat mungojne. Duke qene ripertypes , stomaku perbehet nga kater dhoma: rumen (80%), omasum , retikul dhe aboma . Ciliatet dhe bakteriet e rumenit fermenton komplekset e celuloze duke e kthyer ne acide yndyrore, te cilat me pas thithen permes murit te stomakut. Bovidet kane nje zorre te gjate te vogel ; gjatesia e zorreve te lopeve eshte 29?49 m . Temperatura e trupit luhatet gjate dites; per shembull, tek dhite temperatura mund te ndryshoje pak nga afro 37 ° C ne mengjes heret ne 40 ° C pasdite. Temperatura rregullohet permes djersitjes tek bagetite. Mushkeria e djathte , e perbere nga kater deri ne pese lobe , eshte rreth 1.5 here me e madhe se e majta, e cila ka tre lobe.
[3]
[20]
Shumica e bovideve kane 30 deri ne 32 dhembe. Ndersa incizuesit e siperm mungojne, kaninat e siperme ose zvogelohen ose mungojne. Ne vend te incizuesve te siperm, bovidet kane nje shtrese indi te trashe dhe te ashper, te quajtur jastek dhembesh , i cili siguron nje siperfaqe per te kapur barin dhe gjethet. Ata jane hypsodont dhe selenodont , qe nga molaret dhe premolaret jane te uleta kurorezuar dhe forme hene. Incizoret me te ulet dhe kanellat projektojne perpara. Incizoret percillen nga nje hendek i gjate pa dhembe, i njohur si diastema . Shumica e anetareve te familjes jane barngrenes , por shumica e duikereve jane gjitheckangrenes .
[43]
Ashtu si ripertypesit e tjere, bovidet kane stomake me kater koka, te cilat i lejojne ata te tretin materialin bimor, sic eshte bari , qe nuk mund te perdoret nga shume kafshe te tjera. Ruminantet (dhe disa te tjere si
kanguret
,
lepujt
dhe
termitet
) jane ne gjendje te perdorin
mikroorganizma
qe jetojne ne zorret e tyre per te shkaterruar celulozen nga fermentimi .
[3]
 Antilopat e zeze indiane
Antilopat e zeze indiane
 Gaiali
Gaiali
Bovidet kane metoda te ndryshme te organizimit shoqeror dhe sjelljes shoqerore, te cilat klasifikohen ne sjellje te vetmuar dhe gregare. Me tej, keto lloje secila mund te ndahet ne sjellje territoriale dhe jokritike. Bovidet e vegjel si klipspringeri dhe
oribi
ne pergjithesi jane te vetmuar dhe territorial.
[44]
Ata mbajne territore te vogla ne te cilat anetaret e tjere te specieve nuk lejohen te hyjne. Keto antilopa formojne cifte
monogame
. Shume lloje te tilla si
dik-diku
perdorin sekrecione feromone nga gjendrat preorbital dhe nganjehere jashteshqitje, si dhe, per te shenuar territoret e tyre. Pasardhesit shperndahen ne kohen e
adoleshences
dhe meshkujt duhet te fitojne territore para bashkimit.Tragelafi eshte i vetmi bovid qe eshte edhe i vetmuar dhe jo territorial. Kjo antilope veshtire se tregon agresivitetin, dhe ka tendence te izolohet ose te formoje tufa te lirshme, megjithese ne nje habitat te favorshem, disa tragelaf mund te gjenden mjaft afer njeri-tjetrit.
[45]
Duke perjashtuar cefalofinatet (
duikeret
), tragelafinat (antilopat me brire spirale) dhe neotraginat, shumica e bovideve afrikane jane territorial. Meshkujt detyrohen te shperndahen kur arrijne pjekurine seksuale dhe duhet te formojne territoret e tyre, ndersa femrat nuk u kerkohet ta bejne kete. Meshkujt qe nuk mbajne territore formojne tufat universitare. Konkurrenca zhvillohet midis meshkujve per te fituar mbizoterim, dhe luftimet kane tendence te jene me rigoroze ne sezonet e kufizuara te ndeshjeve . Me perjashtim te meshkujve migratore, meshkujt zakonisht mbajne te njejtin territor gjate gjithe jetes se tyre. Disa individe meshkuj, te njohur si "meshkuj satelitore", mund te lejohen ne territoret e meshkujve te tjere dhe duhet te presin derisa mashkulli kryesor te plaket, ne menyre qe ata te fitojne territorin e tij.
[46]
Sezoni i dukjes eshte ku meshkujt mblidhen se bashku dhe shfaqin konkurrues ndaj bashkeshorteve te mundshem, dihet se ekziston ne mesin e topive dhe kobeve .
[47]
Tragelafet, bagetite, delet dhe dhite jane gregare dhe jo territoriale. Ne keto specie, meshkujt duhet te fitojne dominim absolut mbi te gjithe meshkujt e tjere, dhe luftimet nuk kufizohen vetem ne territore. Meshkujt, pra, kalojne vite ne rritjen e trupit.
[26]
 Antilopa afrikane blu duke luftuar per mbizoterim
Antilopa afrikane blu duke luftuar per mbizoterim
Shumica e bovideve jane
specie ditore
, megjithese disa te tilla si bualli jane perjashtime. Aktiviteti shoqeror dhe ushqyerja zakonisht arrijne kulmin gjate agimit dhe muzgut. Lopet zakonisht pushojne para agimit, gjate mesdites dhe pas erresires. Gershetimi zakonisht behet duke lepire me gjuhen. Rrallehere antilopat rrokullisen ne balte ose pluhur.
Antilopat afrikane
dhe buallicat zakonisht shtrien ne balte, ndersa topit ferkojne koken dhe briret e tyre ne balte dhe me pas i shpartallojne ato mbi trupat e tyre. Bovidet perdorin forma te ndryshme te komunikimit vokal, nuhates dhe te prekshem. Keto perfshijne qendrime te ndryshme te qafes, kokes, brireve, flokeve, kembeve dhe vesheve per te percjelle eksitim seksual, gjendje emocionale ose alarmi. Bovidet zakonisht qendrojne te palevizshem, me koke te larte dhe me veshtrim te ngulitur, kur ndiejne rrezik. Disa si impala,
kudu
dhe elandet madje mund te kercejne ne lartesi disa metra. Bovidet mund te zhurmojne ose grinden per te paralajmeruar te tjeret dhe paralajmeruar grabitqaret. Bovidet te tille si gazelat kercejne ne pergjigje te grabitqareve, duke bere kercime te larta ne kembet e ngurta, duke treguar sinqerisht te dy se grabitqari eshte pare, dhe se individi qe kercen eshte i forte dhe nuk ia vlen te ndiqet.
[48]
 Kercim nga nje Springbok i ri ,sinjale ndershmerie ndaj grabitqaret si gepardet se eshte nje pershtatje dhe te shpejte individual, nuk ia vlen perpjekje per te kapur.
Kercim nga nje Springbok i ri ,sinjale ndershmerie ndaj grabitqaret si gepardet se eshte nje pershtatje dhe te shpejte individual, nuk ia vlen perpjekje per te kapur.
Ne sezonin e ciftezimit, meshkujt qe lutojne per t'ua bere te njohur pranine e tyre grave. Buajt e myshqeve ulerin gjate luftimeve mashkull me mashkull, dhe
saigat
meshkuj forcojne ajrin permes hundeve, duke prodhuar nje zhurme per te penguar meshkujt rivale dhe per te terhequr femrat. Nenat perdorin gjithashtu komunikim vokal per te gjetur vicat e tyre nese ndahen. Gjate luftimeve mbi mbizoterimin, meshkujt kane tendence te shfaqin veten ne nje pozicion te ngritur me nje surrat te nivelit te larte.
[49]
[50]
Teknikat e luftimit ndryshojne ne mesin e familjeve dhe gjithashtu varen nga ndertimi i tyre. Ndersa lufta kongoni ne gjunje, te tjeret zakonisht luftojne ne te kater anet. Gazelet e madhesive te ndryshme perdorin metoda te ndryshme luftimi. Meshkujt e dhive dhe deleve qendrojne drejt dhe perplasen me njeri-tjetrin poshte.Antilopat afrikane perdor koken e fuqishme te kokes ne perleshje agresive. Nese briret ngaterrohen, kundershtaret levizin ne menyre rrethore per t'i zhbllokuar ato. Buajt e myshqeve perplasen me njeri-tjetrin me shpejtesi te larte. Si rregull, vetem dy gjymtyre te njetrajtshme dhe te nivelit te mbrojtjes perfshihen ne nje lufte, e cila ka per qellim te percaktoje eprorin e te dyjave. Individet qe jane dukshem inferiore ndaj te tjereve do te largohen me shume sesa te luftojne; per shembull, meshkujt e papjekur nuk luftojne me demat e pjekur. Ne pergjithesi, bovidet drejtojne sulmet e tyre ne koken e kundershtarit dhe jo ne trupin e tij. Briret ne forme S, te tilla si ato te impales, kane seksione te ndryshme qe ndihmojne ne mbajtjen dhe goditjen me thike. Luftimet serioze qe cojne ne demtim jane te rralla.
[26]
[49]
[51]
 Bovidet jane barngrenes, ushqehen me bar, gjeth dhe produkte bimore.
Bovidet jane barngrenes, ushqehen me bar, gjeth dhe produkte bimore.
Shumica e gjedheve ushqehen dhe piqen ne menyre alternative gjate gjithe dites. Ndersa ato qe ushqehen me ushqim te perqendruar dhe treten ne interval te shkurter, ushqyesit e zjarrit zgjasin intervale me te gjata. Vetem specie te vogla si duikeri ushqehen disa ore gjate dites ose nates. Zakonet e te ushqyerit kane te bejne me madhesine e trupit; ndersa bovidet e vegjel foragjeren ne nje habitat te dendur dhe te mbyllur, speciet me te medha ushqehen me bimesi me
fibra
te larte ne kullota te hapura. Nenfamiljet shfaqin strategji te ndryshme te te ushqyerit. Ndersa speciet Bovina kullosin gjeresisht ne bar te fresket dhe foragjere te shperndare, speciet Cephalophinae (me perjashtim te
Sylvicapra
) kryesisht konsumojne
fruta
. Speciet Reduncinae dhe Hippotraginae varen nga burimet e paqendrueshme te ushqimit, por keto te fundit jane pershtatur posacerisht ne zonat e thata. Anetaret e Caprinae, duke qene ushqyes fleksibel, ushqejne edhe ne zona me produktivitet te ulet. Fiset Alcelaphini, Hippotragini, dhe Reduncini kane permasa te larta te monokoteve ne dietat e tyre. Perkundrazi, Tragelaphini dhe Neotragini (me perjashtim te
Ourebia
) ushqehen gjeresisht me
lule
.
[52]
Asnje marredhenie e dukshme nuk ekziston midis madhesise se trupit dhe konsumit te monocoteve.
[53]
 Qengji prane nenes se saj
Qengji prane nenes se saj
Shumica e bovideve jane poligjen. Ne disa specie, individet jane monogame, duke rezultuar ne agresion minimale mashkull-mashkull dhe zgjedhje te zvogeluar per madhesine e trupit ne meshkuj. Keshtu, dimorfizmi seksual pothuajse mungon. Femrat mund te jene pak me te medha se meshkujt, ndoshta per shkak te
konkurrences
midis femrave per blerjen e territoreve.
[54]
[55]
Ky eshte rasti tek duikeret dhe bovidet e tjere te vegjel. Koha e marre per arritjen e pjekurise seksuale nga secili seks ndryshon gjeresisht midis bovideve.
Pjekuria seksuale
madje mund te paraprihet ose te ndjeke ciftezimin. Per shembull, meshkujt e impales, edhe pse te pjekur seksualisht nga nje vit, mund te ciftezohen vetem pas moshes kater vjec.
[56]
Perkundrazi
delet barbare
femra mund te lindin pasardhes edhe para se te fitojne pjekurine seksuale.
[57]
Vonesa ne pjekurine seksuale tek meshkujve eshte me e dukshme ne speciet seksuale me dimensione seksuale, ndoshta per shkak te konkurrences midis meshkujve. Per shembull, femrat e antilopes afrikane blu behen te afta te riprodhohen brenda nje ose dy vitesh nga lindja, ndersa meshkujt behen te pjekur vetem kur behen kater vjec.
[25]
Te gjithe bovidet bashkohen te pakten nje here ne vit, dhe speciet me te vogla mund te bashkohen edhe dy here.
Stinet
e ciftezimit ndodhin zakonisht gjate muajve me shi per shumicen e bovideve. Si e tille, shumimi mund te arrije dy here ne rajonet ekuatoriale. Delet dhe dhite shfaqin sezonalitet te mrekullueshem te riprodhimit, ne percaktimin e te cilit cikli vjetor i fotoperiodit ditor luan nje rol kryesor. Faktore te tjere qe kane nje ndikim te rendesishem ne kete cikel perfshijne temperaturen e rrethines, statusin ushqyes, nderveprimet shoqerore, daten e ndarjes dhe periudhen e laktacionit. Nje studim i ketij fenomeni arriti ne perfundimin se dhite dhe delet jane mbareshtues te dites se shkurter . Miftezimi ne shumicen e racave te deleve fillon ne vere ose ne fillim te vjeshtes.
[58]
Miftezimi tek delet preket gjithashtu nga melatonin, qe perparon fillimin e sezonit te mbareshtimit;
[59]
dhe tiroksina , qe perfundon sezonin e mbareshtimit.
[60]
Estrusi
zgjat me se shumti nje dite ne gjedhe, me perjashtim te bovineve dhe tragelafineve. Me perjashtim te kongonit dhe topit, te gjithe bovidet mund te zbulojne estrus tek femra duke testuar urinen duke perdorur organin vomeronasal . Pasi mashkulli te sigurohet se femra eshte ne estrus, ai fillon shfaqjet e miqesise; keto shfaqje ndryshojne shume nga marshimet e hollesishme midis specieve gregari deri te shuplaka e zjarrte e organeve gjenitale femerore midis specieve te vetmuara. Femrat, fillimisht jo pranuese, ne fund shoqerohen me mashkullin qe ka arritur mbizoterimin ndaj te tjereve. Receptiviteti shprehet me leje per montim nga mashkulli dhe duke lene anash bishtin nga femra. Perpilimi ne pergjithesi zgjat disa
sekonda
.
[26]
[54]
Periudha e shtatzanise
ndryshon ne mes te bovideve ? ndersa shtatezania e duikerit varion nga 120 deri ne 150 dite, shtatezania e buallit afrikan varion nga 300 ne 330 dite. Zakonisht, lind nje pasardhes i vetem (binjaket jane me pak te shpeshte) dhe ai eshte ne gjendje te qendroje dhe te drejtohet vete brenda nje ore nga lindja. Ne speciet monogame, meshkujt ndihmojne ne mbrojtjen e te rinjve te tyre, por nuk eshte keshtu ne speciet poligjene. Shumica e vicave te porsalindur mbeten te fshehura per nje jave deri ne dy muaj, te ushqyer rregullisht nga nenat e tyre. Ne disa lloje te bovideve, foshnjat fillojne te ndjekin nenat e tyre menjehere ose brenda disa ditesh, si tek impalat. Bovid te ndryshme kane strategji te ndryshme per mbrojtjen e te miturve. Per shembull, ndersa nenat me te egra mbrojne vetem te rinjte e tyre, buallicat shfaqin mbrojtje kolektive. Largimi i gjirit mund te ndodhe qe deri dy muaj (si tek antilopat mbreterore) ose ne fund te nje viti (si ne
bualli i myshkut
).
[54]
[55]
Shumica e bovideve te eger jetojne per 10 deri ne 15 vjet. Speciet me te medha kane tendence te jetojne me gjate; per shembull,
bizoni amerikan
mund te jetoje deri ne 25 vjet dhe gurii deri ne 30 vjet. Jetegjatesia mesatare e individeve shtepiak eshte gati dhjete vjet. Per shembull, dhite kane nje jetegjatesi mesatare prej 12 vjet. Zakonisht meshkujt, kryesisht ne specie poligjen, kane jetegjatesi me te shkurter se femrat. Kjo mund t'i atribuohet disa arsye: shperndarje e hershme e meshkujve te rinj, luftime agresive mashkull-mashkull, prekshmeri ndaj grabitqeve (vecanerisht kur meshkujt jane me pak te shkathet, si kudut), dhe kequshqyerja (duke qene ne madhesi te madhe, mashkulli ka ushqye te larte te kerkesave te cilat mund te mos plotesohen).
[61]
[62]
Richard Despard Estes
sugjeroi qe femrat imitojne karakteristikat sekondare seksuale te mashkullit si briret per te mbrojtur pasardhesit e tyre nga meshkujt mbizoterues. Kjo vecori duket se eshte zgjedhur fuqimisht per te parandaluar vdekshmerine e burrave dhe raportet e pabalancuara te seksit per shkak te sulmeve nga meshkujt agresive dhe shperndarjes se detyruar te meshkujve te rinj gjate adoleshences .
[63]
 Elandi ndodhet ne kullotat e Afrikes.
Elandi ndodhet ne kullotat e Afrikes.
Shumica e specieve te larmishme te bovideve ndodhin ne Afrike. Perqendrimi maksimal eshte ne savanat e Afrikes Lindore. Ne varesi te zakoneve te tyre te ushqyerjes, disa lloje kane rrezatuar ne nje pjese te madhe toke, dhe keshtu verehen disa ndryshime ne morfologjine e dhembeve dhe te gjymtyreve. Duikeret banojne ne pyjet ekuatoriale te shiut, sitatunga ndodhen afer kenetave, kullotat e elandeve banojne, springboku dhe oriksi ndodhen ne
shkretetira
, bongo dhe anoa jetojne ne pyje te dendura, dhe dhite malore dhe takin jetojne ne lartesi te larta. Disa specie bovidesh ndodhen edhe ne Evrope, Azi dhe Ameriken e Veriut. Delet dhe dhite gjenden kryesisht ne Euroazi, megjithese delet barbare dhe stampeket perbejne nje pjese te faunes afrikane. Bualli i myshkut kufizohet ne
tundren
arktike . Disa lloje te bovideve jane zbutur nga njerezit.
Zbutja
i dhive dhe deleve filloi 10 mije vjet me pare, ndersa lopet ishin zbutur rreth 7.5 mije vjet me pare.
[3]
[54]
 Zebut ne
Mumbai
Zebut ne
Mumbai
Zbutje e bovideve ka kontribuar per te zhvendosur varesine e qenieve njerezore nga gjueti dheperparimin e bujqesise. Bovidae perfshin tre nga pese barngrenes te medhenj shtepiak, perdorimi i te cilave eshte perhapur jashte kufijve te tyre origjinal, gjegjesisht bagetite, delet dhe dhite; te gjithe jane nga Euroazia, dhe tani gjenden ne te gjithe boten. Dy speciet e tjera jane
kali
dhe
derri
. Bovidet te tjere te medhejn qe jane zbutur, por qe mbeten brenda vargjet e paraardhesve te tyre te egra jane buallica ,
jaku i zbutur
(nga
jaku i eger
),
Zebu
(nga
auroku indian
),
gaiali
(nga
gauri
) dhe
lopa Bali
(nga
bantengu
).
[54]
Disa antilopat jane zbutur perfshire oriksin , adaksit , elandet dhe konglonket bubal te zhdukur . Orikset e
Egjiptit te Lashte
, adakset dhe kongloket bubal pershkruhen ne mure te gdhendura.
Deshmia me e hershme e zbutjes se bovideve eshte qe nga viti 8000 para Krishtit, duke sugjeruar qe procesi filloi ne
Qipro
dhe
pellgun e Eufratit
.
[64]
 Gezofi Merino eshte me i vleresuari, me shkelqim dhe butesi te madhe.
Gezofi Merino eshte me i vleresuari, me shkelqim dhe butesi te madhe.
Produktet e bulmetit
te tilla si
qumeshti
,
gjalpi
,
gjiza
,
kos
,
dhalla
dhe
djathi
prodhohen kryesisht nga lopet, megjithese qumeshti i deleve, dhise, jakut dhe buallices perdoret gjithashtu ne disa pjese te botes dhe per produktet e shijshme. Per shembull, qumeshti i buallices perdoret per te bere
mocarela
ne
Itali
dhe embelsire gulab jamun ne
Indi
,
[65]
ndersa qumeshti i deleve perdoret per te bere djathe blu Roquefort ne
France
.
[66]
Mishi i vicit eshte nje burim i shkelqyeshem i zinkut, selenit, fosforit, hekurit dhe vitamines B.
[67]
Mishi i bizonit eshte me i ulet ne yndyre dhe kolesterol sesa vici, por ka nje permbajtje me te larte te proteinave.
[68]
Lekura e bovideve eshte e ashper dhe te qendrueshme, me avantazhin shtese se ajo mund te behet ne lekure te trashesi te ndryshme - nga lekure e bute veshje me lekure kepuce. Ndersa dhia dhe lekura e lopes kane nje larmi te gjere te perdorimit, lekura e deleve eshte e pershtatshme vetem per qellime te veshjeve.
[69]
Leshi nga delja e Merino eshte me i miri dhe me i vlefshmi . Leshi i Merino eshte 3-5 i gjate dhe shume i bute. Leshi i trashe, duke qene i qendrueshem dhe rezistent ndaj pilulave , perdoren per te bere rroba te ashpra
[70]
dhe qilima.
 Gote-brir i bere nga Brynjolfur Jonsson e Skarð, Islande , 1598
Gote-brir i bere nga Brynjolfur Jonsson e Skarð, Islande , 1598
Vakti
i kockave eshte nje pleh i rendesishem i pasur me kalcium, fosfor dhe azot , i efektshem ne heqjen e aciditetit te tokes .
[71]
Briret e bovideve jane perdorur si anije te pijshem qe nga lashtesia.
[72]
Bovidae kane paraqitur ne histori qe te pakten nga koha e fabulave te Aesopit nga
Greqia e Lashte
rreth 600 pes. Perrallat nga Aesop perfshijne
Lopen dhe Dele
n ,
Frogun dhe kaun
, dhe
Ujku dhe Qengji
.
[73]
Krijesa mitologjike Kimera, e pershkruar si nje
luan
, me koken e nje dhie qe buronte nga shpina, dhe nje bisht qe mund te mbaronte me koken e gjarprit, ishte nje nga pasardhesit e Typhon dhe Echidna dhe nje moter e perbindesh te tille si Cerberus dhe Hydra Lernaean .
[74]
 |
Wikimedia Commons has media related to
Bovidae
.
|
-
- ^
Stampa:MSW3 Artiodactyla
- ^
"Bovidae"
.
Merriam-Webster online dictionary
. Marre me
7 tetor
2014
.
- ^
a
b
c
d
e
Gomez, W.; Patterson, T. A.; Swinton, J.; Berini, J.
"Bovidae: antelopes, cattle, gazelles, goats, sheep, and relatives"
.
Animal Diversity Web
. University of Michigan Museum of Zoology
. Marre me
7 tetor
2014
.
- ^
Gatesy, J.; Amato, G.; Vrba, E.; Schaller, G. (1997). "A cladistic analysis of mitochondrial ribosomal DNA from the Bovidae".
Molecular Phylogenetics and Evolution
.
7
(3): 303?19.
doi
:
10.1006/mpev.1997.0402
.
PMID
9187090
.
- ^
a
b
c
d
Fernandez, M. H.; Vrba, E. S. (2005).
"A complete estimate of the phylogenetic relationships in Ruminantia: a dated species-level supertree of the extant ruminants"
.
Biological Reviews
.
80
(2): 269?302.
doi
:
10.1017/S1464793104006670
.
PMID
15921052
.
S2CID
29939520
.
- ^
Harrison, T. (2011).
Paleontology and Geology of Laetoli Human Evolution in Context
. Dordrecht: Springer. fq.
363
?465.
ISBN
978-9048-199-624
.
- ^
Demiguel, D.; Sanchez, I. M.; Alba, D. M.; Galindo, J.; Robles, J. M.; Moya-Sola, S. (2012). "First evidence of Azanza and Morales, 1994 (Ruminantia, Bovidae), in the Miocene of the Valles-Penedes Basin (Spain)".
Journal of Vertebrate Paleontology
.
32
(6): 1457?62.
doi
:
10.1080/02724634.2012.696082
.
S2CID
86664298
.
- ^
Harrison, T. (1997).
Neogene Paleontology of the Manonga Valley, Tanzania : A Window into the Evolutionary History of East Africa
. New York: Plenum Press. fq. 113.
ISBN
978-0-306-45471-4
.
- ^
Vrba, E. S.; Schaller, G. (2000).
Antelopes, Deer, and Relatives : Fossil Record, Behavioral Ecology, Systematics, and Conservation
. New Haven: Yale University Press.
ISBN
978-0300-081-428
.
- ^
Stampa:MSW3 Artiodactyla
- ^
Stampa:MSW3 Artiodactyla
- ^
a
b
Savage, R.J.G.; Long, M.R. (1986).
Mammal Evolution: an illustrated guide
. New York: Facts on File. fq.
232?5
.
ISBN
978-0-8160-1194-0
.
- ^
a
b
c
Prothero, D. R.; Schoch, R. M. (2002).
Horns, Tusks, and Flippers : the Evolution of Hoofed Mammals
. Baltimore: Johns Hopkins University Press. fq. 87?90.
ISBN
978-0-8018-7135-1
.
- ^
Hassanin, D.; Douzery, E.J. (1999).
"The tribal radiation of the family Bovidae (Artiodactyla) and the evolution of the mitochondrial cytochrome
b
gene"
(PDF)
.
Molecular Phylogenetics and Evolution
.
13
(2): 227?43.
doi
:
10.1006/mpev.1999.0619
.
PMID
10603253
. Arkivuar nga
origjinali
(PDF)
me 2011-07-20.
- ^
a
b
c
d
e
Gilbert, W. H.; Asfaw, B. (2008).
Homo Erectus : Pleistocene Evidence from the Middle Awash, Ethiopia
. Berkeley: University of California Press. fq. 45?84.
ISBN
978-0-520-25120-5
.
- ^
F. Bibi (gusht 2013).
"A multi-calibrated mitochondrial phylogeny of extant Bovidae (Artiodactyla, Ruminantia) and the importance of the fossil record to systematics"
.
BMC Evolutionary Biology
.
13
: 166.
doi
:
10.1186/1471-2148-13-16
.
PMC
3607928
.
PMID
23339550
.
Mirembajtja CS1: DOI i lire i pashenjuar (
lidhja
)
- ^
{{cite journal|last1=Geraads|first1=D.|last2=El Boughabi|first2=S.|last3=Zouhri|first3=S.|title=A new caprin bovid (Mammalia) from the late Miocene of Morocco|journal=Palaeontologica Africana|date=2012|issue=47|pages=19? === Mioceni i hershem === Mioceni i mesem shenoi perhapjen e bovideve ne Kine dhe nenkontinentin Indian. Sipas Vrba, rrezatimi i nenfamiljes Alcelaphinae filloi ne pjesen e fundit te Miocenit te mesem. Fiset Caprinae ndoshta ndryshuan ne Miocenin e mesme te hershme. Caprini u shfaq ne Miocenin e mesme, dhe duket se jane zevendesuar nga bovidet dhe cervidet e tjere te Euroazis.<ref name=kingdon1989>
Kingdon, J. (1989).
East African Mammals: An Atlas of Evolution in Africa (Volume III, Part C)
. Chicago: University of Chicago press. fq. 1?33.
ISBN
978-0-226-43724-8
.
- ^
Stanley, S. M.; Eldredge, N. (1984). "Evolutionary Pattern and Process in the Sister-Group Alcelaphini-Aepycerotini (Mammalia: Bovidae)".
Living Fossils
. Springer. fq. 62?79.
ISBN
978-1461-382-737
.
- ^
Vrba, E. S.; Burckle, L. H.; Partridge, T. C.; Denton, G. H. (1995).
Paleoclimate and Evolution, with Emphasis on Human Origins
. New Haven: Yale University Press. fq. 24?5.
ISBN
978-0300-063-486
.
- ^
a
b
c
Walton, D.W. (1989).
Fauna of Australia (Volume 1B)
. Canberra: Australian Government Publication Service. fq. 1?14.
ISBN
978-0644-060-561
.
- ^
Hassanin, A. (2015).
"Systematics and Phylogeny of Cattle"
. permbledhur nga Garrick, D.; Ruvinsky, A. (red.).
The Genetics of Cattle
(bot. Second). Oxfordshire, Boston: Cabi. fq. 1?18.
ISBN
9781780642215
.
- ^
Huffman, B.
"Royal antelope"
.
Ultimate Ungulate
. Arkivuar nga
origjinali
me 16 dhjetor 2014
. Marre me
12 dhjetor
2020
.
- ^
Hildyard, A. (2001).
Endangered Wildlife and Plants of the World
. New York: Marshall Cavendish. fq. 769?70.
ISBN
978-0-7614-7200-1
.
- ^
Stampa:EOL
- ^
a
b
Lundrigan, B.; Bidlingmeyer, J. (2000).
"
Connochaetes gnou
: black wildebeest"
.
Animal Diversity Web
. University of Michigan
. Marre me
2013-08-21
.
- ^
a
b
c
d
e
Estes, R. D. (2004).
The Behavior Guide to African Mammals : Including Hoofed Mammals, Carnivores, Primates
(bot. 4th). Berkeley: University of California Press. fq.
7?25
.
ISBN
978-0-520-08085-0
.
- ^
Krausman, P.R.; Casey, A.L. (2012).
"
Addax nasomaculatus
"
.
Mammalian Species
.
807
: 1?4.
doi
:
10.1644/807.1
.
- ^
a
b
Caro, Tim
(2009).
"Contrasting colouration in terrestrial mammals"
.
Philosophical Transactions of the Royal Society B
.
364
(1516): 537?548.
doi
:
10.1098/rstb.2008.0221
.
PMC
2674080
.
PMID
18990666
.
- ^
Cott, H. B.
(1940).
Adaptive Coloration in Animals
. London: Methuen. fq.
88
and plate 25.
- ^
Kiltie, R.A. (janar 1998). "Countershading: Universally deceptive or deceptively universal?".
Trends in Ecology & Evolution
.
3
(1): 21?23.
doi
:
10.1016/0169-5347(88)90079-1
.
PMID
21227055
.
- ^
Cott, H. B.
(1940).
Adaptive Coloration in Animals
. London: Methuen. fq.
53
.
- ^
Leslie, D. M.; Sharma, K. (25 shtator 2009).
"Tetracerus quadricornis (Artiodactyla: Bovidae)"
.
Mammalian Species
.
843
: 1?11.
doi
:
10.1644/843.1
.
- ^
E.C., Mungall (2007).
Exotic Animal Field Guide : Nonnative Hoofed Mammals in the United States
(bot. 1st). College Station: Texas A&M University Press. fq. 197.
ISBN
978-1-58544-555-4
.
- ^
American Livestock Breeds Conservancy
(2009).
"Jacob Sheep"
. Pittsboro, North Carolina: American Livestock Breeds Conservancy. Arkivuar nga
origjinali
me 10 gusht 2011
. Marre me
2011-05-05
.
- ^
Bibi, F.; Bukhsianidze, M.; Gentry, A.; Geraads, D.; Kostopoulos, D.; Vrba, E. (2009).
"The fossil record and evolution of Bovidae: state of the field"
.
Palaeontologia Electronica
.
12
(3): 10A.
- ^
Gatesy, J.; Yelon, D.; DeSalle, R.; Vrba, E. (1992).
"Phylogeny of the Bovidae (Artiodactyla, Mammalia), based on mitochondrial ribosomal DNA sequences"
.
Molecular Biology and Evolution
.
9
(3): 433?446.
doi
:
10.1093/oxfordjournals.molbev.a040734
.
PMID
1584013
.
- ^
Lundrigan, B. (1996).
"Morphology of horns and fighting behavior in the family Bovidae"
.
Journal of Mammalogy
.
77
(2): 462?75.
doi
:
10.2307/1382822
.
JSTOR
1382822
.
- ^
Caro, T. M.; Graham, C. M.; Stoner, C. J.; Flores, M. M. (2003).
"Correlates of horn and antler shape in bovids and cervids"
.
Behavioral Ecology and Sociobiology
.
55
(1): 32?41.
doi
:
10.1007/s00265-003-0672-6
.
S2CID
12030618
.
- ^
a
b
Bro-Jørgensen, J. (2007). "The intensity of sexual selection predicts weapon size in male bovids".
Evolution
.
61
(6): 1316?1326.
doi
:
10.1111/j.1558-5646.2007.00111.x
.
PMID
17542842
.
S2CID
24278541
.
- ^
Ezenwa, V. (2008). "Horns honestly advertise parasite infection in male and female African buffalo".
Animal Behaviour
.
75
(6): 2013?21.
doi
:
10.1016/j.anbehav.2007.12.013
.
S2CID
49240459
.
- ^
Stankowich, T.; Caro, T. (2009).
"Evolution of weaponry in female bovids"
.
Proceedings of the Royal Society B: Biological Sciences
.
276
(1677): 4329?34.
doi
:
10.1098/rspb.2009.1256
.
PMC
2817105
.
PMID
19759035
.
- ^
Packer, C. (1983). "Sexual Dimorphism: The Horns of African Antelopes".
Science
.
221
(4616): 1191?3.
Bibcode
:
1983Sci...221.1191P
.
doi
:
10.1126/science.221.4616.1191
.
PMID
17811523
.
S2CID
37000507
.
- ^
Janis, C.; Jarman, P. (1984). Macdonald, D. (red.).
The Encyclopedia of Mammals
. New York: Facts on File. fq.
498?9
.
ISBN
978-0-87196-871-5
.
- ^
Wyatt, T. D. (2003).
Pheromones and Animal Behaviour: Communication by Smell and Taste
. Cambridge: Cambridge University Press. fq.
97
.
ISBN
978-0-521-48526-5
.
- ^
Ciszek, D.
"Bushbuck"
.
Animal Diversity Web
. University of Michigan Museum of Zoology
. Marre me
28 tetor
2014
.
- ^
T. L., Newell.
"Waterbuck"
.
Animal Diversity Web
. University of Michigan Museum of Zoology
. Marre me
28 tetor
2014
.
- ^
Lott, Dale F. (1991).
Intraspecific Variation in the Social Systems of Wild Vertebrates
. Cambridge University Press. fq.
37
.
ISBN
978-0521370240
.
- ^
Bigalke, R.C. (1972).
"Observations on the behaviour and feeding habits of the springbok
Antidorcas marsupialis
"
.
Zoologica Africana
.
7
(1): 333?359.
doi
:
10.1080/00445096.1972.11447448
.
- ^
a
b
Parker, S.P. (1990).
Grzimek's Encyclopedia of Mammals (Volume 5)
(bot. 1st). New York: McGraw-Hill Publishing. fq.
288?324, 338?9, 354?5, 432?3, 444?5, 460?1, 482?3
.
ISBN
9780079095084
.
- ^
Czaplewski, N. J.; Ryan, J. M.; Vaughan, T. A. (2011).
Mammalogy
(bot. 5th). Sudbury: Jones and Bartlett Publishers.
ISBN
9780763762995
.
- ^
Post, E.; Forchhammer, M. C (korrik 2008).
"Climate change reduces reproductive success of an Arctic herbivore through trophic mismatch"
.
Philosophical Transactions of the Royal Society B: Biological Sciences
.
363
(1501): 2367?2373.
doi
:
10.1098/rstb.2007.2207
.
PMC
2606787
.
PMID
18006410
.
- ^
Gagnon, M.; Chew, A.E. (maj 2000).
"Dietary preferences in extant African Bovidae"
.
Journal of Mammalogy
.
81
(2): 490?511.
doi
:
10.1644/1545-1542(2000)081<0490:DPIEAB>2.0.CO;2
.
- ^
Sponheimer, M.; Lee-Thorp, J.A.; DeRuiter, D.J.; Smith, J.M.; van der Merwe, N.J.; Reed, K.; Grant, C.C.; Ayliffe, L.K.; Robinson, T.F. (2003).
"Diets of Southern African Bovidae: Stable Isotope Evidence"
.
Journal of Mammalogy
.
84
(2): 471?9.
doi
:
10.1644/1545-1542(2003)084<0471:DOSABS>2.0.CO;2
.
- ^
a
b
c
d
e
Feldhamer, George A.; Drickamer, Lee C.; Vessey, Stephen H.; Merritt, Joseph F.; Krajewski, Carey (2007).
Mammalogy: Adaptation, Diversity, Ecology
. Johns Hopkins University Press. fq. 519?522.
ISBN
978-0-8018-8695-9
.
- ^
a
b
Krebs, J.R.; Davies, N.B. (1997).
Behavioural Ecology: An Evolutionary Approach
(bot. 4th). Wiley-Blackwell.
ISBN
9780865427310
.
- ^
Estes, R. D. (2004).
The Behavior Guide to African Mammals : Including Hoofed Mammals, Carnivores, Primates
(bot. 4th). Berkeley: University of California Press. fq.
158?66
.
ISBN
978-0-520-08085-0
.
- ^
Gray, Gary G.; Simpson, C. David (1980). "Ammotragus lervia".
Mammalian Species
.
1980
(144): 1?7.
doi
:
10.2307/3504009
.
JSTOR
3504009
.
- ^
Rosa, H.J.D.; Bryant, M.J. (2003). "Seasonality of reproduction in sheep".
Small Ruminant Research
.
48
(3): 155?71.
doi
:
10.1016/S0921-4488(03)00038-5
.
- ^
Chemineau, P.; Pelletier, J.; Guerin, Y.; Colas, G.; Ravault, J.P.; Toure, G.; Almeida, G.; Thimonier, J.; Ortavant, R. (1988).
"Photoperiodic and melatonin treatments for the control of seasonal reproduction in sheep and goats"
.
Reproduction, Nutrition, Development
.
28
(2B): 409?22.
doi
:
10.1051/rnd:19880307
.
PMID
3413339
.
- ^
Prendergast, B. J.; Mosinger, B.; Kolattukudy, P. E.; Nelson, R. J. (2002).
"Hypothalamic gene expression in reproductively photoresponsive and photorefractory Siberian hamsters"
.
Proceedings of the National Academy of Sciences
.
99
(25): 16291?6.
Bibcode
:
2002PNAS...9916291P
.
doi
:
10.1073/pnas.232490799
.
PMC
138604
.
PMID
12456888
.
- ^
Owen-Smith, N. (1993).
"Comparative mortality rates of male and female kudus: the costs of sexual size dimorphism"
.
Journal of Animal Ecology
.
62
(3): 428?40.
doi
:
10.2307/5192
.
JSTOR
5192
.
- ^
Toigo, C.; Gaillard, J.M. (2003). "Causes of Sex-Biased Adult Survival in Ungulates: Sexual Size Dimorphism, Mating Tactic or Environment Harshness?".
Oikos
.
101
(2): 376?84.
doi
:
10.1034/j.1600-0706.2003.12073.x
.
- ^
Estes, R.D. (1991). "The significance of horns and other male secondary sexual characters in female bovids".
Applied Animal Behaviour Science
.
29
(1?4): 403?51.
doi
:
10.1016/0168-1591(91)90264-X
.
- ^
Zeder, M. A. (2006).
Documenting Domestication : New Genetic and Archaeological Paradigms
. Berkeley, California: University of California Press. fq.
317
.
ISBN
978-0520246386
.
- ^
Phelan, Benjamin; Phelan, Benjamin (24 korrik 2013).
"Others' Milk"
. Slate.com
. Marre me
10 tetor
2014
.
- ^
Hughes, Tom; Hughes, Meredith Sayles (2005).
Gastronomie!: Food Museums and Heritage Sites of France
. Bunker Hill Publishing. fq.
19
.
ISBN
978-1-59373-029-1
.
- ^
"Beef, lean organic"
. WHFoods. 18 tetor 2004. Arkivuar nga
origjinali
me 2 prill 2015
. Marre me
1 prill
2015
.
- ^
"| National Bison Association"
. Bisoncentral.com.
Arkivuar
nga origjinali me 20 janar 2011
. Marre me
1 prill
2015
.
- ^
Veldmeijer, A. J.; Harris, S. (2014).
Why Leather?: The Material and Cultural Dimensions of Leather
. Sidestone Press. fq. 31?6.
ISBN
9789088902611
.
- ^
"Merino Sheep in Australia"
. Arkivuar nga
origjinali
me 2006-11-05
. Marre me
1 prill
2015
.
- ^
Kolay, A. K. (2007).
Manures and fertilizers
. New Delhi: Atlantic Publications. fq. 98.
ISBN
978-8126908103
.
- ^
Vi, E. (1975).
"Germanic glass drinking horns"
.
Journal of Glass Studies
.
17
: 74?87.
- ^
"Aesop's Fables"
.
Aesop's Fables
. Arkivuar nga
origjinali
me 16 tetor 2014
. Marre me
10 tetor
2014
.
- ^
Peck.
"Entry:Chimaera"
. Marre me
31 mars
2015
.
</ref Delet, sinonim i dhise ne mitologjine kineze, eshte kafsha e tete e zodiakut kinez dhe nje simbol i devotshmerise filiale.<ref name=eberhard>
Eberhard, W.
A Dictionary of Chinese Symbols: Hidden Symbols in Chinese Life and Thought
. London: Routledge.
ISBN
978-0-415-00228-8
.