 Осамен грабливец:
бела мечка
се храни со брадеста фока што ?а убила.
Осамен грабливец:
бела мечка
се храни со брадеста фока што ?а убила.
 Соци?ални грабливци : мравките од месо соработуваат за да се хранат со цикада многу поголема од нив.
Соци?ални грабливци : мравките од месо соработуваат за да се хранат со цикада многу поголема од нив.
Грабливост
или
предаци?а
? биолошки однос каде што еден организам,
грабливецот
(
предатор
), убива и ?аде друг организам, неговиот
плен
. Тоа е едно од семе?ството на вообичаени однесува?а за хране?е кое вклучува
паразитизам
и
микрограбливост
(кои обично не го убиваат дома?инот) и паразитоидизам (што секогаш го прави, на кра?от). Се разликува од
чисте?ето
на мртвиот плен, иако многу грабливци исто така чистат; се преклопува со
тревопасните животни
, биде??и грабливците на семи?ата и деструктивните фру?адци се грабливци.
Грабливците можат активно да бараат или да го гонат пленот или да го чекаат, честопати скриен. Кога е откриен плен, грабливецот проценува дали да го нападне. Ова може да вклучува заседа или потера пред грабнува?е, понекогаш по следе?е на пленот. Ако нападот е успешен, грабливецот го убива пленот, ги отстранува сите не?адливи делови како лушпата или боцките и го ?аде.
Грабливците се приспособени и често се високо специ?ализирани за лов, со акутни сетила како што се
видот
, слухот или мирисот . Многу грабливи
животни
, и
’рбетници
и
безрбетници
, имаат остри кан?и или вилици за да го фатат, убиваат и исечат пленот. Други адаптации вклучуваат способност за крие?е и агресивна мимикри?а кои ?а подобруваат ефикасноста на ловот.
Грабливоста има мо?ен селективен ефект врз пленот, а пленот развива противграбливски приспособува?а, како што се
предупредувачко обо?ува?е
, алармни повици и други сигнали, камуфлажа,
мимикри?а
на добро одбранети видови и одбранбени боцки и хемикалии. Понекогаш грабливецот и пленот се нао?аат во еволутивна трка во вооружува?е, циклус на адаптации и контра-адаптации. Грабливоста е главен двигател на
еволуци?ата
барем од
камбрискиот
период.
 Па?аковите оси ги парализираат и на кра?от ги убиваат своите дома?ини, но се сметаат за паразитоиди, а не за грабливци.
Па?аковите оси ги парализираат и на кра?от ги убиваат своите дома?ини, но се сметаат за паразитоиди, а не за грабливци.
На на?основно ниво, грабливците убиваат и ?адат други организми. Сепак, концептот на грабливост е широк, различно дефиниран во различни контексти и вклучува широк спектар на методи на хране?е; а некои врски што резултираат со смрт на пленот генерално не се нарекуваат грабливост. Паразитоид, како што е ихнеумонската оса, ги положува ?а?цата во или на сво?от дома?ин; ?а?цата се отвораат во ларви, кои го ?адат дома?инот и то? неизбежно умира. Зоолозите генерално го нарекуваат ова форма на
паразитизам
, иако конвенционално се смета дека паразитите не ги убиваат своите дома?ини. Може да се дефинира дека грабливецот се разликува од паразитоидот по тоа што има многу плен, заробен во текот на неговиот живот, каде што ларвата на паразитоид има само еден, или барем има обезбедено снабдува?е со храна за него само во една прилика.
[1]
[2]
 Поврзаност на грабливоста со други стратегии за хране?е
Поврзаност на грабливоста со други стратегии за хране?е
Има и други тешки и гранични случаи.
Микрограбливците
се мали животни кои, како и грабливците, целосно се хранат со други организми; тие вклучуваат
болви
и
комарци
кои консумираат крв од живи животни и
вошки
кои консумираат сок од живи растени?а. Ме?утоа, биде??и тие обично не ги убиваат своите дома?ини, тие сега често се сметаат за паразити.
[3]
[4]
Животните кои
пасат
на
фитопланктон
или душеци од микроби се грабливци, биде??и ги консумираат и убиваат нивните организми за храна; но тревопасните животни кои прелистуваат лис?а не се, биде??и нивните растени?а кои им служат како храна обично го преживуваат нападот.
[5]
Кога животните ?адат семи?а (
грабливост
на семи?а или
зрна
) или ?а?ца (
грабливост
на ?а?ца), тие консумираат цели живи организми, што по дефиници?а ги прави грабливци.
[6]
[7]
[8]
Чистачите
, организми кои ?адат само организми прона?дени ве?е мртви, не се грабливци, но многу грабливци како
шакалот
и
хиената
чистат кога ?е се укаже прилика.
[5]
[9]
[10]
Ме?у безрбетниците,
соци?алните оси
(жолти ?акни) се и ловци и чистачи на други инсекти.
[11]
</img>
Грабливост на семи?а:
глушец
?аде семи?а
Додека примерите на грабливци ме?у цицачите и птиците се добро познати,
[12]
грабливците може да се на?дат во широк опсег на таксони вклучува??и членконоги. Тие се вообичаени ме?у инсектите, вклучително и мантиди, вилински ко?чи?а, врвки и скорпии . Ка? некои видови, како што е алдерот, само ларвите се грабливи (возрасните не ?адат). Па?аците се грабливи, како и другите копнени безрбетници како што се
скорпиите
; стоногалки; некои грини, полжави и голтки;
нематоди
; и планарни црви.
[13]
Во морските средини, пове?ето
книдари
(на пр.
медузи
, хидроиди ),
ктенофора
(желе со чешел),
ехинодерми
(на пример,
морски ?везди
,
морски ежови
, песочни долари и
морски краставици
) и
рамни црви
се грабливи. Ме?у
раковите
, ?астозите,
раковите
, ракчи?ата и шталите се грабливци,
[14]
а за возврат раковите се плен на речиси сите
главонози
(вклучува??и
октоподи
, лиг?и и сипи ).
[15]
 Парамециум
, граблив цили?ат, ко? се храни со
бактерии
Парамециум
, граблив цили?ат, ко? се храни со
бактерии
Прабливоста на семи?ата е ограничена на цицачи, птици и инсекти, но го има во речиси сите копнени екосистеми.
[6]
[8]
Грабливоста на ?а?цата ги вклучува и специ?алните грабливци на ?а?ца, како што се некои
змии
од
колобриди
, и генералистите, како што се лисиците и ?азовците, кои опортунистички земаат ?а?ца кога ?е ги на?дат.
[16]
[17]
[18]
Некои растени?а, како стомното растение, стапицата за муви на Венера и слаткото, се
месо?адни и консумираат инсекти
.
[12]
Методите на грабеж од страна на растени?ата варираат во голема мера, но често вклучуваат замка за храна, механичка стимулаци?а и електрични импулси за на кра?от да го фатат и консумираат пленот.
[19]
Некои месо?адни габи фа?аат
нематоди
користе??и или активни стапици во форма на стегачки прстени или пасивни стапици со лепливи структури.
[20]
Многу видови на
протозои
(
еукариоти
) и
бактерии
(
прокариоти
) плен на други микроорганизми; начинот на хране?е е очигледно антички и еволуирал многу пати во двете групи.
[12]
[21]
[22]
Поме?у слатководните и морските
зоопланктони
, без разлика дали се едноклеточни или пове?еклеточни, грабливото пасе?е на
фитопланктонот
и помалите зоопланктони е вообичаено и се сре?ава во многу видови нанофлагелати, динофлагелати, цили?ати,
ротифери
, разновидна палета на меровапланктонски групи и животински групи на ракови, имено копеподи и кладоцерани.
[23]
 Основен циклус на потрага по храна за грабливец, со одредени вари?ации
[24]
Основен циклус на потрага по храна за грабливец, со одредени вари?ации
[24]
За да се нахрани, грабливецот мора да го бара, да го следи и да го убие сво?от плен. Овие де?ства формираат циклус на потрага по храна.
[25]
[26]
Грабливецот мора да одлучи каде да бара плен врз основа на неговата географска местоположба; и штом ?е на?де плен, мора да процени дали да го следи или да чека подобар избор. Ако избере потера, неговите физички способности го одредуваат начинот на потера (на пример, заседа или потера).
[27]
[28]
Откако ?е го фати пленот, можеби ?е треба да троши енерги?а и за
ракува?е со
него (на пр., убива?е, отстранува?е на секо?а школка или боцки и голта?е).
[24]
[25]
Грабливците имаат избор на начини на пребарува?е кои се движат од
седе?е и чека?е
до
активно
или
широко бара?е храна
.
[24]
[29]
[30]
[31]
Методот на седе?е и чека?е е на?соодветен ако пленот е густ и подвижен, а грабливецот има ниски бара?а за енерги?а.
[29]
Широкото бара?е храна троши пове?е енерги?а и се користи кога пленот е седентарен или ретко распространет.
[27]
[29]
Постои континуитет на режими на пребарува?е со интервали поме?у периодите на движе?е кои се движат од секунди до месеци. А?кулите, сончевите рипчи?а,
инсекто?адните
птици и
итрините
речиси секогаш се движат додека па?аците кои градат мрежа, водните безрбетници, богомолките и ветрушките ретко се движат. Поме?у, краставиците и другите кра?брежни птици, слатководните риби, вклучително и слатките, и ларвите на кокцинелидните
бубачки (бубамари)
, наизменично се движат ме?у активно пребарува?е и скенира?е на околината.
[29]
 Албатросот со црни ве?и редовно лета стотици километри низ речиси празниот океан за да на?де парчи?а храна.
Албатросот со црни ве?и редовно лета стотици километри низ речиси празниот океан за да на?де парчи?а храна.
Распределбите на пленот честопати се собираат на едно место, а грабливците реагираат со бара?е каде што пленот е на?густо распределен, а потоа бараат во закрпи.
[24]
Онаму каде што храната се нао?а во групи, како што се ретки дождовни риби во речиси празен океан, фазата на пребарува?е бара од грабливецот да патува значително време и да потроши значителна количина на енерги?а, за да го лоцира секое парче храна.
[29]
На пример, црно-ве?естиот албатрос редовно прави летови за потрага по храна во опсег од околу 700 километри (430 мил?и), до максимален опсег на потрага по храна од 3000 километри (1860 мил?и) за размножува?е на птици кои собираат храна за своите млади.
[б 1]
[32]
Со статичен плен, некои грабливци можат да научат соодветни локации на групи и да се вра?аат ка? нив во интервали за да се хранат.
[29]
Оптималната стратеги?а за потрага по храна за пребарува?е е моделирана со помош на теоремата за маргинална вредност.
[33]
Моделите за пребарува?е често се по?авуваат случа?но. Една таква е прошетка Леви, ко?а има тенденци?а да вклучува кластери од кратки чекори со повремени долги чекори. Добро одговара на однесува?ето на широк спектар на организми, вклучува??и бактерии, пчели, а?кули и човечки ловци-собирачи.
[34]
[35]
 Бубамарите со седум точки
избираат растени?а со добар квалитет за нивниот плен на
вошки
.
Бубамарите со седум точки
избираат растени?а со добар квалитет за нивниот плен на
вошки
.
Откако ?е на?де плен, грабливецот мора да одлучи дали да го следи или да продолжи да го бара. Одлуката зависи од вклучените трошоци и придобивки. Птицата ко?а бара инсекти троши многу време во потрага, но нивното фа?а?е и ?аде?е е брзо и лесно, така што ефикасната стратеги?а за птицата е да го ?аде секо? вкусен инсект што ?е го на?де. Спротивно на тоа, грабливец како лав или сокол лесно го нао?а сво?от плен, но за да го фати бара многу напор. Во то? случа?, грабливецот е поселективен.
[27]
Еден од факторите што треба да се земе предвид е големината. Премалиот плен можеби не е вреден за маката поради количината на енерги?а што ?а дава. Премногу голем и можеби е премногу тешко да се улови. На пример, мантидата фа?а плен со своите предни нозе и тие се оптимизирани за граба?е плен со одредена големина. Мантидите не сакаат да напа?аат плен што е далеку од таа големина. Постои позитивна корелаци?а поме?у големината на грабливецот и неговиот плен.
[27]
Грабливецот исто така може да процени група и да одлучи дали да помине време во потрага по плен во неа.
[24]
Ова може да вклучи одредено знае?е за преференциите на пленот; на пример,
бубамарите
можат да изберат парче вегетаци?а погодна за нивниот плен на
вошки
.
[36]
За да го фатат пленот, грабливците имаат спектар на начини на потера кои се движат од отворена потера (грабливост по
потера
) до ненадеен удар на плен во близина (
гребливост од заседа
).
[12]
[24]
[37]
Друга стратеги?а поме?у заседата и потера е
балистичкото пресретнува?е
, каде што грабливецот го наб?удува и предвидува движе?ето на пленот и потоа го започнува сво?от напад соодветно.
[38]
Грабливците кои чекаат од заседа се месо?адни животни кои го фа?аат пленот скришум или изненадувачки. Ка? животните, грабливецот од заседа се одликува со тоа што грабливецот ?а скенира околината од скриена положба додека не се забележи плен, а потоа брзо извршува фиксен изненаден напад.
[38]
[39]
Грабливците од заседа на ’рбетниците вклучуваат жаби, риби како што се ангелската а?кула, северната штука и источната жаба риба.
[38]
[40]
[41]
[42]
Ме?у многуте грабливци од заседа безрбетници се па?аците од врата и па?аците од австралиски рак на копно и ракчи?ата богомолка во морето.
[39]
[43]
[44]
Грабливците од заседа често градат дупка во ко?а ?е се сокри?ат, подобрува??и го прикрива?ето по цена на намалува?е на нивното видно поле. Некои грабливци од заседа користат и мамки за да привлечат плен во опсегот на удар.
[38]
Движе?ето за фа?а?е треба да биде брзо за да се зароби пленот, има??и предвид дека нападот не може да се измени откако ?е се започне.
[38]
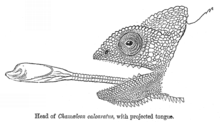 Камелеонот
го напа?а пленот испушта??и го ?азикот.
Камелеонот
го напа?а пленот испушта??и го ?азикот.
Балистичкото пресретнува?е е стратеги?а каде што грабливецот го наб?удува движе?ето на пленот, го предвидува неговото движе?е, изработува патека за пресретнува?е и потоа го напа?а пленот на таа патека. Ова се разликува од грабливката од заседа по тоа што грабливецот го прилагодува сво?от напад според тоа како се движи пленот.
[38]
Балистичкото пресретнува?е вклучува краток период за планира?е, дава??и му можност на пленот да избега. Некои жаби чекаат додека змиите не почнат да удираат пред да скокнат, намалува??и го времето на располага?е на зми?ата да го рекалибрира не?зиниот напад и максимизира??и го аголното прилагодува?е што зми?ата треба да го направи за да ?а пресретне жабата во реално време.
[38]
Балистичките грабливци вклучуваат инсекти како што се вилинските ко?чи?а и ’рбетници како што се стрелците (напа?аат со млаз вода),
камелеони
(напа?аат со ?азикот) и некои
колубридни змии
.
[38]
</img>
Китови со грпка
се хранилки за лансира?е, филтрираат ил?адници
крили
од морската вода и ги голтаат живи.
Во потерата на грабливците, грабливците бркаат плен што бега. Ако пленот бега во права лини?а, фа?а?ето зависи само од тоа дали грабливецот е побрз од пленот.
[38]
Ако пленот маневрира врте??и сe додека бега, грабливецот мора да реагира во реално време за да пресмета и следи нова патека за пресретнува?е, како на пример со паралелна навигаци?а, додека се затвора врз пленот.
[38]
Многу грабливци за потера користат камуфлажа за да му при?дат на пленот што е можно поблиску незабележано (
демнат
) пред да започнат со потера.
[38]
Грабливците на потера вклучуваат копнени цицачи како што се лу?е, африкански диви кучи?а, забележани хиени и волци; морски грабливци како што се делфини, орки и многу грабливи риби, како што е туната;
[45]
[46]
грабливи птици (грабливци) како соколи; и инсекти како вилински ко?чи?а .
[47]
Екстремна форма на потера е истра?ниот или упорниот лов, во ко? грабливецот го уморува пленот следе??и го на голема далечина, понекогаш и со часови. Ово? метод го користат ловци-собирачи и
каниди
како што се
африканските диви кучи?а
и домашните песови. Африканското диво куче е екстремен упорен грабливец, ко? го заморува индивидуалниот плен следе??и го многу мил?и со релативно мала брзина.
[48]
Специ?ализирана форма на грабеж за потера е хране?ето на балените китови . Овие многу големи морски грабливци се хранат со
планктон
, особено крил, нурка??и и активно плива??и во концентрации на планктон, а потоа земаат огромна голтка вода и ?а филтрираат низ нивните пердувести балински плочи.
[49]
[50]
Грабливците од потера може да бидат соци?ални, како лавот и волкот што ловат во групи или сами.
[2]
Штом грабливецот ?е го фати пленот, мора да се справи со него: многу внимателно ако пленот е опасен за ?аде?е, како на пример ако поседува остри или отровни боцки, како ка? многу риби плен. Некои сомови како што се Ictaluridae имаат боцки на грбот (дорзален) и стомак (пекторален) кои се блокираат во исправена положба; додека сом тепа кога ?е биде фатен, тие би можеле да ?а проби?ат устата на грабливецот, можеби фатално. Некои птици што ?адат риба, како што е
оспреот
, ?а избегнуваат опасноста од боцки со тоа што го кинат пленот пред да го ?адат.
[51]
Осамен наспроти соци?ален грабливец
[
уреди
|
уреди извор
]
In social predation, a group of predators cooperates to kill prey. This makes it possible to kill creatures larger than those they could overpower singly; for example,
hyenas
, and
wolves
collaborate to catch and kill herbivores as large as buffalo, and lions even hunt elephants.
[52]
[53]
[54]
It can also make prey more readily available through strategies like flushing of prey and herding it into a smaller area. For example, when mixed flocks of birds forage, the birds in front flush out insects that are caught by the birds behind. Spinner dolphins form a circle around a school of fish and move inwards, concentrating the fish by a factor of 200.
[55]
By hunting socially
chimpanzees
can catch colobus monkeys that would readily escape an individual hunter, while cooperating Harris hawks can trap rabbits.
[52]
[56]
 Волци
, соци?ални грабливци, соработуваат за да ловат и убиваат
бизони
.
Волци
, соци?ални грабливци, соработуваат за да ловат и убиваат
бизони
.
Грабливците од различни видови понекогаш соработуваат за да фатат плен. Во
коралните гребени
, кога рибите како груперот и коралната пастрмка ?е забележат недостапен плен за нив, тие им сигнализираат на ?иновските ?агули, наполеоновите и
октоподите
. Овие грабливци можат да пристапат до мали пукнатини и да го исфрлат пленот.
[57]
[58]
Познато е дека
китовите уби?ци
им помагаат на китовите да ловат китови .
Соци?алниот лов им овозможува на грабливците да се справат со поширок опсег на плен, но со ризик од конкуренци?а за заробената храна. Осамените грабливци имаат пове?е шанси да го изедат тоа што ?е го фатат, по цена на зголемена потрошувачка на енерги?а за да го фатат и зголемен ризик дека пленот ?е избега.
[59]
[60]
Грабливците од заседа често се осамени за да го намалат ризикот самите да станат плен.
[61]
Од 245 копнени членови на
Carnivora
(групата ко?а ги вклучува мачките, кучи?ата и мечките), 177 се самосто?ни ловци; и 35 од 37
диви мачки
се сами,
[62]
вклучува??и ги и пумата и гепардот.
[2]
[59]
Сепак, осамената пума им дозволува на другите пуми да учествуваат во убиството,
[63]
и
ко?отот
може да биде или осамен или соци?ален.
[64]
Други осамени грабливци ги вклучуваат северната штука,
[65]
волч?и па?аци и сите ил?адници видови осамени оси ме?у членконогите,
[66]
и многу
микроорганизми
и
зоопланктон
.
[21]
[67]
Под притисок на
природната селекци?а
, грабливците еволуирале различни физички
адаптации
за открива?е, фа?а?е, убива?е и варе?е на пленот. Тие вклучуваат брзина, агилност, способност за крие?е, остри сетила, кан?и, заби, филтри и соодветни дигестивни системи.
[68]
За открива?е на плен, грабливците имаат добро развиен
вид
, мирис или
слух
.
[12]
Разновидните грабливци како бувовите и па?аците кои скокаат имаат очи свртени нанапред, обезбедува??и точен двогледен вид на релативно тесно видно поле, додека грабливките често имаат помалку акутен сеопфатен вид. Животните како што се лисиците можат да го мирисаат сво?от плен дури и кога е скриен под 2 стапки (60 cм) снег или зем?а. Многу грабливци имаат акутен слух, а некои како што се
лил?аците
со ехолокаци?а ловат исклучиво со активна или пасивна употреба на звук.
[69]
Грабливци, вклучува??и ги големите мачки,
птици грабливки
и мравки имаат мо?ни вилици, остри заби или кан?и кои ги користат за да го зграпчат и уби?ат сво?от плен. Некои грабливци како што се
змиите
и птиците што ?адат риби како
чап?ите
и корморани го голтаат сво?от плен цел; некои змии можат да ги откачат вилиците за да им овозможат да голтаат голем плен, додека птиците што ?адат риби имаат долги клунови налик на коп?е што ги користат за да го прободат и фатат брзо-движечкиот и лизгав плен.
[69]
Рибите и другите грабливци развиле способност да ги дробат или отвораат оклопните школки на мекотелите.
[70]
Многу грабливци се мо?но изградени и можат да фатат и убиваат животни поголеми од себе; ова важи исто толку за малите грабливци како што се
мравките
и
итрините
, како и за големите и видливо мускулести месо?ади како
пумата
и
лавот
.
[2]
[69]
[71]
</img>
Грабливост по големина:
лавица
напа?а
бивол од Ке?п
, пове?е од дво?но поголема од не?зината тежина. Лавовите можат да нападнат многу поголем плен, вклучително и слоновите, но тоа го прават многу поретко.
Грабливците често се високо специ?ализирани во нивната исхрана и лове?е; на пример,
евроазискиот рис
лови само мали копитари.
[72]
Други, како што се
леопардите
, се поопортунистички генералисти, плен на на?малку 100 видови.
[73]
[74]
Специ?алистите може да бидат многу прилагодени да го фа?аат сво?от претпочитан плен, додека генералистите може подобро да се префрлат на друг плен кога претпочитаната цел е ретка. Кога пленот има згрчена (нерамномерна) распределба, се предвидува дека оптималната стратеги?а за грабливецот ?е биде поспеци?ализирана биде??и пленот е повпечатлив и може побрзо да се на?де;
[75]
се чини дека ова е точно за грабливците на неподвижен плен, но е сомнително со мобилниот плен.
[76]
Со грабливците кои се избираат според големината, грабливците избираат плен со одредена големина.
[77]
Големиот плен може да биде проблематичен за грабливецот, додека малиот плен може да биде тешко да се на?де и во секо? случа? да обезбеди помала награда. Ова доведе до корелаци?а поме?у големината на грабливците и нивниот плен. Големината може да делува и како засолниште за голем плен. На пример, возрасните слонови се релативно безбедни од грабеж од лавови, но малолетниците се ранливи.
[78]
Членовите на
семе?ството на мачки
, како што се
снежниот леопард
(висорамнини без дрв?а),
тигарот
(тревни рамнини, мочуришта од трска),
оцелото
(шума), рибарска мачка (гради покра? водата) и
лавот
(отворени рамнини) се камуфлирани со бо?а и нарушувачки шари кои одговараат на нивните живеалишта.
[79]
Во агресивната мимикри?а, одредени грабливци, вклучува??и инсекти и риби, користат бо?а и однесува?е за да привлечат плен. Женките Фотурис светулки, на пример, ги копираат светлосните сигнали на други видови, со што привлекуваат машки светулки, кои ги фа?аат и ?адат. Цветните богомолки се грабливци од заседа; камуфлирани како цве?и?а, како орхидеи, тие привлекуваат плен и го грабнуваат кога е доволно блиску. Жабарите се екстремно добро камуфлирани и активно го намамуваат сво?от плен да се приближи со помош на еска, мамка на кра?от од додатокот на главата како прачка, со ко? нежно мавтаат за да имитираат мало животно, голта??и го пленот со исклучително брзо движе?е. кога е во домет.
Многу помали грабливци, како што е медузата, користат отров за да го покорат сво?от плен,
[80]
а отровот исто така може да помогне во варе?ето (како што е случа?от со змиите ?вечарка и некои
па?аци
).
[81]
[82]
Мермерната морска зми?а ко?а се приспособила на грабнува?е на ?а?ца има атрофирани отровни жлезди, а генот за не?зиниот токсин од три прсти содржи
мутаци?а
(со брише?е на два
нуклеотида
) што ги деактивира. Овие промени се об?аснуваат со фактот дека неговиот плен не треба да се покори.
[83]
 Електричен зрак (Torpediniformes ) што ?а покажува локаци?ата на електричниот орган и електроцитите наредени во него
Електричен зрак (Torpediniformes ) што ?а покажува локаци?ата на електричниот орган и електроцитите наредени во него
Неколку групи на грабливи риби имаат способност да детектираат, следат, а понекогаш, како во електричниот зрак, да го онеспособат сво?от плен со
насетува?е и генерира?е на електрични поли?а
.
[84]
[85]
[86]
Електричниот орган е изведен од модифицирано нервно или мускулно ткиво.
[87]
Физиолошките адаптации на грабливката ?а вклучуваат способноста на грабливите бактерии да го варат сложениот полимер на
пептидогликан
од
клеточните ?идови
на бактериите што ги пленат.
[22]
Месо?адните ’рбетници од сите пет главни класи (риби, водоземци, рептили, птици и цицачи) имаат пониски релативни стапки на транспорт на ше?ер до
аминокиселини
од тревопасните или сешто?адите, веро?атно затоа што тие добиваат многу аминокиселини од животинските
белковини
во нивната исхрана.
[88]
Противграбливски приспособува?а
[
уреди
|
уреди извор
]
За да се спротивстави на грабливката, пленот еволуирал одбрана за употреба во секо?а фаза од нападот. Тие можат да се обидат да избегнат открива?е, како на пример со употреба на камуфлажа и мимикри?а. Тие можат да откри?ат грабливци и да ги предупредат другите за нивното присуство. Доколку бидат откриени, тие можат да се обидат да избегнат да бидат цел на напад, на пример, со сигнализира?е дека се токсични или непри?атни, со формира?е групи или дека потера би била непрофитабилна. Ако станат цел, тие можат да се обидат да го одвратат нападот со одбранбени средства како што се оклоп, пердуви, невкусност или мобинг; и тие можат да избегнат напад што е во тек со заплашува?е на грабливецот, со преправа?е дека се мртви, фрла?е делови од телото како опашки или едноставно бегство.
 Лил?аците користат ехолокаци?а за да ловат молци но?е.
Лил?аците користат ехолокаци?а за да ловат молци но?е.
Грабливците и пленот се природни непри?атели, а многу од нивните адаптации изгледаат диза?нирани да се спротивстават еден на друг. На пример, лил?аците имаат софистицирани системи за ехолокаци?а за открива?е на инсекти и друг плен, а инсектите развиле разновидни одбранбени средства, вклучително и способност да ги слушаат повиците за ехолокаци?а.
[89]
[90]
Многу грабливци кои трчаат на копно, како што се волците, развиле долги екстремитети како одговор на зголемената брзина на нивниот плен.
[91]
Нивните адаптации се окарактеризирани како еволутивна трка во вооружува?е, пример за
коеволуци?а
на два вида.
[92]
Во поглед на еволуци?ата фокусиран на гените, гените на грабливецот и пленот може да се смета дека се натпреваруваат за телото на пленот.
[92]
Сепак, принципот ?живот-вечера“ на Докинс и Кребс предвидува дека оваа трка во вооружува?е е асиметрична: ако грабливецот не успее да го фати сво?от плен, ?а губи вечерата, додека ако успее, пленот го губи животот.
[92]
 Источната корална зми?а, ко?а и самата е грабливец, е доволно отровна за да ги убие грабливците што ?а напа?аат, па кога ?е ?а избегнат, ова однесува?е мора да се наследи, а не да се научи.
Источната корална зми?а, ко?а и самата е грабливец, е доволно отровна за да ги убие грабливците што ?а напа?аат, па кога ?е ?а избегнат, ова однесува?е мора да се наследи, а не да се научи.
Метафората за трка во вооружува?е подразбира посто?ано ескалира?е на напредок во нападот и одбраната. Сепак, овие адаптации доа?аат со трошок; на пример, подолгите нозе имаат зголемен ризик од крше?е,
[93]
додека специ?ализираниот ?азик на камелеонот, со неговата способност да делува како проектил, е бескорисен за преклопува?е на вода, па камелеонот мора да пие роса од вегетаци?ата.
[94]
Принципот ?живот-вечера“ е критикуван на пове?е основи. Степенот на асиметри?ата во природната селекци?а делумно зависи од наследноста на адаптивните особини.
[94]
Исто така, ако грабливецот изгуби доволно вечери, и то? ?е го загуби животот.
[93]
[94]
Од друга страна, цената на фитнес способноста за дадена изгубена вечера е непредвидлив, биде??и грабливецот може брзо да на?де подобар плен. Покра? тоа, пове?ето грабливци се генералисти, што го намалува вли?анието на дадената адаптаци?а на плен врз грабливецот. Биде??и специ?ализаци?ата е предизвикана од коеволуци?ата грабливец-плен, реткоста на специ?алисти може да значи дека трките во вооружува?е грабливец-плен се ретки.
[94]
Тешко е да се одреди дали дадените адаптации се навистина резултат на коеволуци?ата, каде што адаптаци?ата на пленот доведува до адаптаци?а на грабливец што се спротивставува со понатамошна адаптаци?а на пленот. Алтернативно об?аснува?е е
ескалаци?ата
, каде што грабливците се прилагодуваат на конкурентите, нивните сопствени грабливци или опасниот плен.
[95]
Очигледните приспособува?а на грабливоста, исто така, може да се по?ават од други причини, а потоа да бидат избрани за напад или одбрана. Ка? некои од инсектите плен од лил?аци, слухот еволуирал пред да се по?ават лил?аците и се користел за слуша?е сигнали што се користат за територи?ална одбрана и паре?е.
[89]
Нивниот слух еволуирал како одговор на грабливоста на лил?аците, но единствениот ?асен пример за реципрочна адаптаци?а ка? лил?аците е скришната ехолокаци?а.
[89]
Посиметрична трка во вооружува?е може да се случи кога пленот е опасен, има боцки, пердуви, токсини или отров што може да му наштетат на грабливецот. Грабливецот може да одговори со избегнува?е, што пак ?а поттикнува еволуци?ата на мимикри?ата. Избегнува?ето не е нужно еволутивен одговор биде??и генерално се учи од лошите искуства со пленот. Ме?утоа, кога пленот е способен да го убие грабливецот (како и коралната зми?а со сво?от отров), нема можност за уче?е и избегнува?ето мора да се наследи. Грабливците можат да одговорат и на опасниот плен со контра-адаптации. Во западна Северна Америка, обичната зми?а жартиера развила отпорност на токсинот во кожата на тритонот со груба кожа .
[94]
Грабливците вли?аат на нивните екосистеми не само директно со ?аде?е на сопствениот плен, туку и со индиректни средства, како што е намалува?е на грабливката од други видови или менува?е на однесува?ето на тревопасниот живот во потрага по храна, како што е ефектот на биодиверзитетот на волците врз вегетаци?ата покра? реката или морските видри на шумите со алги. . Ова може да ги об?асни ефектите од динамиката на популаци?ата, како што се циклусите забележани ка? рисот и за?аците со чевли за снег.
[96]
[97]
[98]
Еден начин за класификаци?а на грабливците е според трофично ниво . Месо?адите кои се хранат со
тревопасни животни
се секундарни потрошувачи; нивните грабливци се терци?арни потрошувачи, и така натаму.
[99]
На врвот на ово?
син?ир
на исхрана се на?високите грабливци како што се
лавовите
.
[100]
Сепак, многу грабливци ?адат од пове?е нивоа на син?ирот на исхрана; месо?ад може да ?аде и секундарни и терци?арни потрошувачи.
[101]
Ова значи дека многу грабливци мора да се борат со интраеснафскиот грабливец, каде што другите грабливци ги убиваат и ?адат. На пример,
ко?отите
се натпреваруваат и понекогаш убиваат сиви лисици и бобкети .
[102]
Биолошка разновидност одржана со врвна грабливост
[
уреди
|
уреди извор
]
Грабливците може да го зголемат
биодиверзитетот
на заедниците со тоа што ?е спречат еден вид да стане доминантен. Таквите грабливци се познати како клучни видови и може да имаат големо вли?ание врз рамнотежата на организмите во одреден
екосистем
.
[103]
Воведува?ето или отстранува?ето на ово? грабливец, или промените во неговата густина на населеност, може да имаат драстични каскадни ефекти врз рамнотежата на многу други популации во екосистемот. На пример, животните кои пасат на пасиштата може да спречат еден доминантен вид да преземе.
[104]
 Закрепнува?е на кра?брежна врба во Blacktail Creek,
националниот парк ?елоустоун
, по повторното воведува?е на волци, локалниот
вид клучен камен
и
врвниот грабливец
.
[105]
Лево, во 2002 година; точно, во 2015 година
Закрепнува?е на кра?брежна врба во Blacktail Creek,
националниот парк ?елоустоун
, по повторното воведува?е на волци, локалниот
вид клучен камен
и
врвниот грабливец
.
[105]
Лево, во 2002 година; точно, во 2015 година
Елиминаци?ата на волците од
Националниот парк ?елоустоун
имало големо вли?ание врз трофичката пирамида . Во таа област, волците се и клучни видови и врвни грабливци. Без грабливост, тревопасните животни почнале прекумерно да пасат многу дрвенести видови, што вли?аело на популациите на растени?ата во областа. Покра? тоа, волците често ги спречувале животните да пасат во близина на потоци, заштитува??и ги изворите на храна на дабарите . Отстранува?ето на волците имало директно вли?ание врз популаци?ата на дабарите, биде??и нивното живеалиште станало територи?а за пасе?е. Зголеменото прелистува?е на
врбите
и
четинарите
долж Blacktail Creek поради недостаток на грабливци предизвикало засекува?е на каналот биде??и намалената популаци?а на дабари пове?е не можела да ?а забави водата и да ?а одржува почвата на своето место. На то? начин се покажало дека грабливците се од витално значе?е во екосистемот.
[105]
 Бро? на крзна од за?аци со снежни чевли (
Lepus americanus
) (жолта позадина) и канадски рис (црна лини?а, преден план) продадени на компани?ата Хадсон Бе? од 1845 до 1935 година
Бро? на крзна од за?аци со снежни чевли (
Lepus americanus
) (жолта позадина) и канадски рис (црна лини?а, преден план) продадени на компани?ата Хадсон Бе? од 1845 до 1935 година
Во отсуство на грабливци, популаци?ата на еден вид може да расте експоненци?ално додека не се приближи до носивоста на животната средина.
[106]
Грабливците го ограничуваат растот на пленот и со нивно консумира?е и со менува?е на нивното однесува?е.
[107]
Зголемува?ето или намалува?ето на популаци?ата на плен може да доведе и до зголемува?е или намалува?е на бро?от на грабливци, на пример, преку зголемува?е на бро?от на младенчи?а што ги носат.
Циклични флуктуации се забележани ка? популациите на грабливец и плен, често со поместува?а поме?у циклусите на грабливец и плен. Добро познат пример е оно? на за?акот и
рисот
. Во текот на широк распон на
бореални шуми
во Ал?аска и Канада, популаци?ата на за?аците флуктуира речиси синхронизирано во период од 10 години, а популациите на рисот флуктуираат како одговор на тоа. Ова за прв пат беше видено во историските записи за животни фатени од ловци на крзно за компани?ата Хадсон Бе? пред пове?е од еден век.
[98]
[108]
[109]
[110]
 Циклуси на популаци?ата грабливец - плен во моделот Лотка-Волтера
Циклуси на популаци?ата грабливец - плен во моделот Лотка-Волтера
Едноставен модел на систем со по еден вид на грабливец и плен, равенките Лотка-Волтера, ги предвидува циклусите на популаци?ата.
[111]
Сепак, обидите да се репродуцираат предвидува?ата на ово? модел во лаборатори?а честопати не успеваат; на пример, кога протозоата
Didinium nasutum
се додава на култура ко?а го содржи неговиот плен,
Paramecium caudatum
, вториот често се доведува до исчезнува?е.
[112]
Равенките Лотка-Волтера се потпираат на неколку поедноставувачки претпоставки и тие се структурно нестабилни, што значи дека секо?а промена во равенките може да ?а стабилизира или дестабилизира динамиката.
[113]
[114]
На пример, една претпоставка е дека грабливците имаат линеарен функционален одговор на пленот: стапката на убиства се зголемува пропорционално со стапката на средби. Ако оваа стапка е ограничена со времето поминато за ракува?е со секо? улов, тогаш популаци?ата на плен може да достигне густина над ко?а грабливците не можат да ги контролираат.
[112]
Друга претпоставка е дека секо? поединечен плен е идентичен. Во реалноста, грабливците имаат тенденци?а да избираат млади, слаби и болни поединци, остава??и ?а популаци?ата на плен способна повторно да расте.
[115]
Многу фактори можат да ?а стабилизираат популаци?ата на грабливци и плен.
[116]
Еден пример е присуството на пове?екратни грабливци, особено генералисти кои се привлечени од даден вид плен ако е изобилен и бараат на друго место ако не е.
[116]
Како резултат на тоа, циклусите на популаци?а имаат тенденци?а да се на?дат во северните умерени и
субарктички
екосистеми, биде??и мрежите за храна се поедноставни.
[116]
Системот на за?аци и рисови со чевли за снег е субарктички, но дури и тоа вклучува други грабливци, вклучува??и ко?оти, гошавки и големи рогови бувови, а циклусот е засилен со вари?ации во храната достапна за за?аците.
[116]
Развиени се низа математички модели со релаксира?е на претпоставките направени во моделот Лотка-Волтера; тие на различни начини им дозволуваат на животните да имаат географска дистрибуци?а или да мигрираат ; да има разлики ме?у поединците, како што се
полот
и старосната структура, така што само некои поединци се репродуцираат; да живеат во различна средина, како на пример со менува?е на
годишните времи?а
;
[117]
[118]
и анализира?е на интеракциите на пове?е од само два вида одеднаш. Ваквите модели предвидуваат многу различни и често хаотични динамики на популаци?ата грабливец-плен.
[117]
[119]
Присуството на засолништа, каде што пленот е безбеден од грабливци, може да му овозможи на пленот да одржува поголеми популации, но исто така може да ?а дестабилизира динамиката.
[120]
[121]
[122]
[123]
Предира?ето датира од пред по?авата на општо признатите месо?ади со стотици милиони (можеби мили?арди) години. Грабливоста еволуирала посто?ано ка? различни групи на организми.
[5]
[124]
Подемот на
еукариотските
клетки на околу 2,7 Gya, порастот на пове?еклеточните организми на околу 2 Gya и порастот на мобилните грабливци (околу 600 Mya - 2 Gya, веро?атно околу 1 Gya) се припишуваат на раното грабливо однесува?е, и многу многу рани остатоци покажуваат докази за дупнатини или други ознаки кои им се припишуваат на малите видови грабливци.
[5]
Веро?атно предизвикало големи еволутивни транзиции, вклучува??и го пристигнува?ето на
клетки
,
еукариоти
, сексуална репродукци?а,
пове?еклеточност
, зголемена големина, подвижност (вклучува??и го летот на инсектите
[125]
) и оклопни школки и егзоскелети.
[5]
На?раните грабливци биле микробни организми кои ги голтале или паселе други. Биде??и фосилните записи се лоши, овие први грабливци би можеле да датираат поме?у 1 и над 2,7 Gya (пред мили?арди години)
[5]
Грабливоста видно станала важна непосредно пред
Камбрискиот
период - пред околу 550 милиони години - како што е потврдено од речиси истовремениот разво? на калцификаци?а ка? животните и алгите,
[126]
избегнува??и дупче?е од грабливост. Сепак, грабливците паселе микроорганизми од пред на?малку 1,000 милиони години,
[5]
[127]
[128]
со докази за селективно (наместо случа?но) грабнува?е од слично време.
[129]
Фосилниот запис
демонстрира долга истори?а на интеракции поме?у грабливците и нивниот плен од камбрискиот период па наваму, покажува??и на пример дека некои грабливци дупчеле низ лушпите на мекотелите со
бивалвни
и
гастроподи
, додека други ги ?аделе овие организми со крше?е на нивните лушпи.
[130]
Ме?у камбриските грабливци имало безрбетници како аномалокаридидите со додатоци погодни за граба?е плен, големи сложени очи и вилици направени од тврд матери?ал како оно? во
егзоскелетот
на инсектот.
[131]
Некои од првите
риби што имале вилици
биле оклопните и главно грабливи
плакодерми
од
Силурскиот
до
девонскиот
период, од кои едната, 6 метарската
Dunkleosteus
, се смета за првиот
’рбетник
во светот ?надграбливец“, плен на други грабливци.
[132]
[133]
Инсектите
развиле способност да летаат во раниот
карбон
или доцниот девон, овозможува??и им, ме?у другото, да избегаат од грабливците.
[125]
Ме?у на?големите грабливци кои некогаш живееле биле
тероподните диносауруси
како што е
Тираносаурус
од периодот на
креда
. Тие ловеле тревопасни диносауруси како што се хадросаурусите, цератопсиите и анкилосаурусите.
[134]
-
Камбриската револуци?а на подлогата
виде дека животот на
морското дно
се смени од минимално закопува?е (лево) во разновидна фауна за вкопува?е (десно), веро?атно за да се избегнат новите
камбриски
грабливци.
-
Устата на
аномалокаридидната
Laggania cambria
,
камбриски
безрбетник, веро?атно врвен грабливец
-
Dunkleosteus
,
девонски
плакодерм
, можеби првиот суперграбливец на ’рбетници во светот, реконструкци?а
-
Meganeura monyi
, грабливи
карбонски
инсект
поврзан со
вилинските ко?чи?а
, може да лета за да избега од копнените грабливци. Неговата голема големина, со распон на крил?ата од 65 cм (30 ин), може да го одрази недостатокот на ’рбетници воздушни грабливци во тоа време.
-
 Сан Хантер, Боцвана
Сан Хантер, Боцвана
Лу?ето, како
сешто?ади
, се до одреден степен грабливи,
[135]
користат оруж?е и алатки за лове?е
риби
,
[136]
ловат
и заробуваат животни.
[137]
Тие користат и други грабливи видови како кучи?а, корморани
[138]
и
соколи
за да фатат плен за храна или за спорт.
[139]
Два грабливци со средна големина, кучи?а и мачки, се животните кои на?често се чуваат како
домашни миленици
во западните општества.
[140]
[141]
Лу?ето ловци, вклучително и Сан од ?ужна Африка, користат упорност во ловот, форма на грабеж по потера, каде што гонителот може да биде побавен од пленот, како што е антилопата куду на кратки расто?ани?а, но го следи на пладневна топлина додека не се исцрпи, потера ко?а може да потрае и до пет часа.
[142]
[143]
Во биолошката контрола на штетници, грабливци (и паразитоиди) од природниот опсег на штетници се воведуваат за контрола на популациите, со ризик да предизвикаат непредвидени проблеми. Природните грабливци, под услов да не им штетат на видовите кои не се штетници, се еколошки и одржлив начин за намалува?е на штетите врз посевите и алтернатива на употребата на хемиски агенси како што се
пестицидите
.
[144]
 Капитолинскиот волк ги дои
Ромул и Рем
, митските основачи на
Рим
Капитолинскиот волк ги дои
Ромул и Рем
, митските основачи на
Рим
Во филмот, иде?ата за грабливец како опасен
хуманоиден
непри?ател се користи во
научно-фантастичниот
хорор
акционен филм ?
<i id="mwBJI">Предатор</i> “ од 1987 година и неговите три продолжени?а .
[145]
Застрашувачки грабливец, огромна
бела а?кула
што ?аде човек, е исто така централно место во трилерот на
Стивен Спилберг
од 1974 година ?
Вилици
“.
[146]
Во поези?ата на тема грабливост, може да се истражи свеста на грабливецот, како на пример во
Штука
на
Тед Х?уз
.
[147]
Фразата ?Природа, црвена во забите и кан?ите“ од Алфред, песната на Лорд Тенисон од 1849 година ? In Memoriam AHH “ се толкува како да се однесува на борбата поме?у грабливците и пленот.
[148]
Во митологи?ата и народната басна, грабливците како лисицата и волкот имаат измешана репутаци?а.
[149]
Лисицата била симбол на плодноста во Стара Грци?а, но временски демон во северна Европа и суштество на ?аволот во раното христи?анство; лисицата е претставена како итра, алчна и лукава во басните од
Езоп
наваму.
[149]
Големиот лош волк им е познат на децата во приказните како
Црвенкапа
, но е демонска фигура во исландските Еда саги, каде што волкот Фенрир се по?авува во апокалиптичниот
кра? на светот
.
[149]
Во средниот век, верува?ето се раширило во
врколаците
, лу?ето се трансформирале во волци.
[149]
Во Стариот Рим и Египет, волкот се обожавал, волчицата се по?авува во основачкиот мит за Рим, дое??и ги
Ромул и Рем
.
[149]
Неодамна, во Книгата за
?унглата
на
Ради?ард Киплинг
од 1894 година, Могли е воспитан од глутницата волци.
[149]
Ставовите кон големите грабливци во Северна Америка, како што се волкот,
гризли
и пумата, се префрли?а од непри?ателство или амбивалентност, придружени со активно прогонство, кон позитивни и заштитнички во втората половина на 20 век.
[150]
- Екологи?а на стравот
- Проблем со грабливост
- Превртува?е грабливец-плен
- Ва-Тор
- ↑
A range of 3000 kilometres means a flight distance of at least 6000 kilometres out and back.
- Beauchamp, Guy (2012).
Social predation : how group living benefits predators and prey
. Elsevier.
ISBN
9780124076549
.
- Bell, W. J. (2012).
Searching Behaviour : the behavioural ecology of finding resources
. Springer Netherlands.
ISBN
9789401130981
.
- Caro, Tim
(2005).
Antipredator Defenses in Birds and Mammals
. University of Chicago Press.
ISBN
978-0-226-09436-6
.
- Cott, Hugh B.
(1940).
Adaptive Coloration in Animals
. Methuen.
OCLC
974070031
.
- Jacobs, David Steve; Bastian, Anna (2017).
Predator-prey interactions : co-evolution between bats and their prey
. Springer.
ISBN
9783319324920
.
- Rockwood, Larry L. (2009).
Introduction to population ecology
. John Wiley & Sons. стр. 281.
ISBN
9781444309102
.
- Ruxton, Graeme D.
;
Sherratt, Tom N.
; Speed, Michael P. (2004).
Avoiding attack: the evolutionary ecology of crypsis, warning signals, and mimicry
. Oxford University Press.
ISBN
9780198528593
.
- ↑
Gurr, Geoff M.; Wratten, Stephen D.; Snyder, William E. (2012).
Biodiversity and Insect Pests: Key Issues for Sustainable Management
. John Wiley & Sons. стр. 105.
ISBN
978-1-118-23185-2
.
- ↑
2,0
2,1
2,2
2,3
Lafferty, K. D.; Kuris, A. M. (2002). ?Trophic strategies, animal diversity and body size“.
Trends Ecol. Evol
.
17
(11): 507?513.
doi
:
10.1016/s0169-5347(02)02615-0
.
- ↑
Poulin, Robert; Randhawa, Haseeb S. (February 2015).
?Evolution of parasitism along convergent lines: from ecology to genomics“
.
Parasitology
.
142
(Suppl 1): S6?S15.
doi
:
10.1017/S0031182013001674
.
PMC
4413784
.
PMID
24229807
.
- ↑
Poulin, Robert
(2011). Rollinson, D.; Hay, S. I. (уред.).
The Many Roads to Parasitism: A Tale of Convergence
.
Advances in Parasitology
.
74
. Academic Press. стр. 27?28.
doi
:
10.1016/B978-0-12-385897-9.00001-X
.
ISBN
978-0-12-385897-9
.
PMID
21295676
.
- ↑
5,0
5,1
5,2
5,3
5,4
5,5
5,6
Bengtson, S. (2002). ?Origins and early evolution of predation“. Во Kowalewski, M.; Kelley, P. H. (уред.).
The fossil record of predation. The Paleontological Society Papers 8
(PDF)
. The Paleontological Society. стр. 289?317. Архивирано од
изворникот
(PDF)
на 2008-09-10
. Посетено на
2022-04-26
.
- ↑
6,0
6,1
Janzen, D. H. (1971). ?Seed Predation by Animals“.
Annual Review of Ecology and Systematics
.
2
: 465?492.
doi
:
10.1146/annurev.es.02.110171.002341
.
- ↑
Nilsson, Sven G.; Bjorkman, Christer; Forslund, Par; Hoglund, Jacob (1985). ?Egg predation in forest bird communities on islands and mainland“.
Oecologia
.
66
(4): 511?515.
Bibcode
:
1985Oecol..66..511N
.
doi
:
10.1007/BF00379342
.
PMID
28310791
.
- ↑
8,0
8,1
Hulme, P. E.; Benkman, C. W. (2002). C. M. Herrera and O. Pellmyr (уред.).
Granivory
.
Plant animal Interactions: An Evolutionary Approach
. Blackwell. стр. 132?154.
ISBN
978-0-632-05267-7
.
- ↑
Kane, Adam; Healy, Kevin; Guillerme, Thomas; Ruxton, Graeme D.; Jackson, Andrew L. (2017). ?A recipe for scavenging in vertebrates ? the natural history of a behaviour“.
Ecography
.
40
(2): 324?334.
doi
:
10.1111/ecog.02817
.
- ↑
Kruuk, Hans (1972).
The Spotted Hyena: A Study of Predation and Social Behaviour
. University of California Press. стр. 107?108.
ISBN
978-0226455082
.
- ↑
Schmidt, Justin O. (2009). ?Wasps“.
Wasps - ScienceDirect
.
Encyclopedia of Insects
(Second. изд.). стр. 1049?1052.
doi
:
10.1016/B978-0-12-374144-8.00275-7
.
ISBN
9780123741448
.
- ↑
12,0
12,1
12,2
12,3
12,4
Stevens, Alison N. P. (2010).
?Predation, Herbivory, and Parasitism“
.
Nature Education Knowledge
.
3
(10): 36.
- ↑
?Predators, parasites and parasitoids“
.
Australian Museum
(англиски)
. Посетено на
19 September
2018
.
- ↑
Phelan, Jay (2009).
What Is life? : a guide to biology
(Student. изд.). W.H. Freeman & Co. стр. 432.
ISBN
9781429223188
.
- ↑
Villanueva, Roger; Perricone, Valentina; Fiorito, Graziano (17 August 2017).
?Cephalopods as Predators: A Short Journey among Behavioral Flexibilities, Adaptions, and Feeding Habits“
.
Frontiers in Physiology
.
8
: 598.
doi
:
10.3389/fphys.2017.00598
.
PMC
5563153
.
PMID
28861006
.
- ↑
Hanssen, Sveinn Are; Erikstad, Kjell Einar (2012). ?The long-term consequences of egg predation“.
Behavioral Ecology
.
24
(2): 564?569.
doi
:
10.1093/beheco/ars198
.
- ↑
Pike, David A.; Clark, Rulon W.; Manica, Andrea; Tseng, Hui-Yun; Hsu, Jung-Ya; Huang, Wen-San (2016-02-26).
?Surf and turf: predation by egg-eating snakes has led to the evolution of parental care in a terrestrial lizard“
.
Scientific Reports
.
6
(1): 22207.
Bibcode
:
2016NatSR...622207P
.
doi
:
10.1038/srep22207
.
PMC
4768160
.
PMID
26915464
.
- ↑
Ainsworth, Gill; Calladine, John; Martay, Blaise; Park, Kirsty; Redpath, Steve; Wernham, Chris; Wilson, Mark; Young, Juliette (2017).
Understanding Predation: A review bringing together natural science and local knowledge of recent wild bird population changes and their drivers in Scotland
. Scotland's Moorland Forum. стр. 233?234.
doi
:
10.13140/RG.2.1.1014.6960
.
- ↑
Hedrich, Rainer; Fukushima, Kenji (20 May 2021).
?On the Origin of Carnivory: Molecular Physiology and Evolution of Plants on an Animal Diet“
.
Annual Review of Plant Biology
.
72
(1). annurev?arplant?080620-010429.
doi
:
10.1146/annurev-arplant-080620-010429
.
ISSN
1543-5008
.
PMID
33434053
. Архивирано од
изворникот
на 2022-05-07
. Посетено на
2022-04-26
.
- ↑
Pramer, D. (1964). ?Nematode-trapping fungi“.
Science
.
144
(3617): 382?388.
Bibcode
:
1964Sci...144..382P
.
doi
:
10.1126/science.144.3617.382
.
JSTOR
1713426
.
PMID
14169325
.
- ↑
21,0
21,1
Velicer, Gregory J.; Mendes-Soares, Helena (2007).
?Bacterial predators“
(PDF)
.
Cell
.
19
(2): R55?R56.
doi
:
10.1016/j.cub.2008.10.043
.
PMID
19174136
.
- ↑
22,0
22,1
Jurkevitch, Edouard; Davidov, Yaacov (2006). ?Phylogenetic Diversity and Evolution of Predatory Prokaryotes“.
Predatory Prokaryotes
. Springer. стр.
11
?56.
doi
:
10.1007/7171_052
.
ISBN
978-3-540-38577-6
.
- ↑
Hansen, Per Juel; Bjørnsen, Peter Koefoed; Hansen, Benni Winding (1997). ?Zooplankton grazing and growth: Scaling within the 2-2,-μm body size range“.
Limnology and Oceanography
.
42
(4): 687?704.
Bibcode
:
1997LimOc..42..687H
.
doi
:
10.4319/lo.1997.42.4.0687
.
summarizes findings from many authors.
- ↑
24,0
24,1
24,2
24,3
24,4
24,5
Kramer, Donald L. (2001).
?Foraging behavior“
(PDF)
. Во Fox, C. W.; Roff, D. A.; Fairbairn, D. J. (уред.).
Evolutionary Ecology: Concepts and Case Studies
. Oxford University Press. стр. 232?238.
ISBN
9780198030133
. Архивирано од
изворникот
(PDF)
на 12 July 2018
. Посетено на
20 September
2018
.
- ↑
25,0
25,1
Griffiths, David (November 1980). ?Foraging costs and relative prey size“.
The American Naturalist
.
116
(5): 743?752.
doi
:
10.1086/283666
.
JSTOR
2460632
.
- ↑
Wetzel, Robert G.; Likens, Gene E. (2000).
?Predator-Prey Interactions“
.
Limnological Analyses
. Springer. стр.
257?262
.
doi
:
10.1007/978-1-4757-3250-4_17
.
ISBN
978-1-4419-3186-3
.
- ↑
27,0
27,1
27,2
27,3
Pianka, Eric R. (2011).
Evolutionary ecology
(7 (eBook). изд.). Eric R. Pianka. стр. 78?83.
- ↑
MacArthur, Robert H. (1984). ?The economics of consumer choice“.
Geographical ecology : patterns in the distribution of species
. Princeton University Press. стр. 59?76.
ISBN
9780691023823
.
- ↑
29,0
29,1
29,2
29,3
29,4
29,5
Bell 2012
harvnb error: no target: CITEREFBell2012 (
help
)
- ↑
Eastman, Lucas B.; Thiel, Martin (2015). ?Foraging behavior of crustacean predators and scavengers“. Во Thiel, Martin; Watling, Les (уред.).
Lifestyles and feeding biology
. Oxford University Press. стр. 535?556.
ISBN
9780199797066
.
- ↑
Perry, Gad (January 1999). ?The Evolution of Search Modes: Ecological versus Phylogenetic Perspectives“.
The American Naturalist
.
153
(1): 98?109.
doi
:
10.1086/303145
.
PMID
29578765
.
- ↑
Gremillet, D.; Wilson, R. P.; Wanless, S.; Chater, T. (2000). ?Black-browed albatrosses, international fisheries and the Patagonian Shelf“.
Marine Ecology Progress Series
.
195
: 69?280.
Bibcode
:
2000MEPS..195..269G
.
doi
:
10.3354/meps195269
.
- ↑
Charnov, Eric L. (1976).
?Optimal foraging, the marginal value theorem“
(PDF)
.
Theoretical Population Biology
.
9
(2): 129?136.
doi
:
10.1016/0040-5809(76)90040-x
.
PMID
1273796
.
- ↑
Reynolds, Andy (September 2015). ?Liberating Levy walk research from the shackles of optimal foraging“.
Physics of Life Reviews
.
14
: 59?83.
Bibcode
:
2015PhLRv..14...59R
.
doi
:
10.1016/j.plrev.2015.03.002
.
PMID
25835600
.
- ↑
Buchanan, Mark (5 June 2008). ?Ecological modelling: The mathematical mirror to animal nature“.
Nature
.
453
(7196): 714?716.
doi
:
10.1038/453714a
.
PMID
18528368
.
- ↑
Williams, Amanda C.; Flaxman, Samuel M. (2012). ?Can predators assess the quality of their prey's resource?“.
Animal Behaviour
.
83
(4): 883?890.
doi
:
10.1016/j.anbehav.2012.01.008
.
- ↑
Scharf, Inon; Nulman, Einat; Ovadia, Ofer; Bouskila, Amos (September 2006). ?Efficiency evaluation of two competing foraging modes under different conditions“.
The American Naturalist
.
168
(3): 350?357.
doi
:
10.1086/506921
.
PMID
16947110
.
- ↑
38,00
38,01
38,02
38,03
38,04
38,05
38,06
38,07
38,08
38,09
38,10
Moore, Talia Y.; Biewener, Andrew A. (2015).
?Outrun or Outmaneuver: Predator?Prey Interactions as a Model System for Integrating Biomechanical Studies in a Broader Ecological and Evolutionary Context“
(PDF)
.
Integrative and Comparative Biology
.
55
(6): 1188?97.
doi
:
10.1093/icb/icv074
.
PMID
26117833
.
- ↑
39,0
39,1
deVries, M. S.; Murphy, E. A. K.; Patek S. N. (2012). ?Strike mechanics of an ambush predator: the spearing mantis shrimp“.
Journal of Experimental Biology
.
215
(Pt 24): 4374?4384.
doi
:
10.1242/jeb.075317
.
PMID
23175528
.
- ↑
?Cougar“
.
Hinterland Who's Who
.
Canadian Wildlife Service
and
Canadian Wildlife Federation
. Архивирано од
изворникот
на 18 May 2007
. Посетено на
22 May
2007
.
- ↑
?Pikes (Esocidae)“
(PDF)
. Indiana Division of Fish and Wildlife
. Посетено на
3 September
2018
.
- ↑
Bray, Dianne.
?Eastern Frogfish, Batrachomoeus dubius“
.
Fishes of Australia
. Архивирано од
изворникот
на 14 September 2014
. Посетено на
14 September
2014
.
- ↑
?Trapdoor spiders“
. BBC
. Посетено на
12 December
2014
.
- ↑
?Trapdoor spider“
. Arizona-Sonora Desert Museum. 2014
. Посетено на
12 December
2014
.
- ↑
Gazda, S. K.; Connor, R. C.; Edgar, R. K.; Cox, F. (2005).
?A division of labour with role specialization in group-hunting bottlenose dolphins (Tursiops truncatus) off Cedar Key, Florida“
.
Proceedings of the Royal Society
.
272
(1559): 135?140.
doi
:
10.1098/rspb.2004.2937
.
PMC
1634948
.
PMID
15695203
.
- ↑
Tyus, Harold M. (2011).
Ecology and Conservation of Fishes
. CRC Press. стр. 233.
ISBN
978-1-4398-9759-1
.
- ↑
Combes, S. A.; Salcedo, M. K.; Pandit, M. M.; Iwasaki, J. M. (2013). ?Capture Success and Efficiency of Dragonflies Pursuing Different Types of Prey“.
Integrative and Comparative Biology
.
53
(5): 787?798.
doi
:
10.1093/icb/ict072
.
PMID
23784698
.
- ↑
Hubel, Tatjana Y.; Myatt, Julia P.; Jordan, Neil R.; Dewhirst, Oliver P.; McNutt, J. Weldon; Wilson, Alan M. (2016-03-29).
?Energy cost and return for hunting in African wild dogs“
.
Nature Communications
.
7
: 11034.
doi
:
10.1038/ncomms11034
.
PMC
4820543
.
PMID
27023457
.
Cursorial hunting strategies range from one extreme of transient acceleration, power and speed to the other extreme of persistence and endurance with prey being fatigued to facilitate capture.Dogs and humans are considered to rely on endurance rather than outright speed and manoeuvrability for success when hunting cursorially.
- ↑
Goldbogen, J. A.; Calambokidis, J.; Shadwick, R. E.; Oleson, E. M.; McDonald, M. A.; Hildebrand, J. A. (2006). ?Kinematics of foraging dives and lunge-feeding in fin whales“.
Journal of Experimental Biology
.
209
(7): 1231?1244.
doi
:
10.1242/jeb.02135
.
PMID
16547295
.
- ↑
Sanders, Jon G.; Beichman, Annabel C.; Roman, Joe; Scott, Jarrod J.; Emerson, David; McCarthy, James J.; Girguis, Peter R. (2015).
?Baleen whales host a unique gut microbiome with similarities to both carnivores and herbivores“
.
Nature Communications
.
6
: 8285.
Bibcode
:
2015NatCo...6.8285S
.
doi
:
10.1038/ncomms9285
.
PMC
4595633
.
PMID
26393325
.
- ↑
Forbes, L. Scott (1989). ?Prey Defences and Predator Handling Behaviour: The Dangerous Prey Hypothesis“.
Oikos
.
55
(2): 155?158.
doi
:
10.2307/3565418
.
JSTOR
3565418
.
- ↑
52,0
52,1
Lang, Stephen D. J.; Farine, Damien R. (2017).
?A multidimensional framework for studying social predation strategies“
.
Nature Ecology & Evolution
.
1
(9): 1230?1239.
doi
:
10.1038/s41559-017-0245-0
.
PMID
29046557
.
- ↑
MacNulty, Daniel R.; Tallian, Aimee; Stahler, Daniel R.; Smith, Douglas W. (2014-11-12). Sueur, Cedric (уред.).
?Influence of Group Size on the Success of Wolves Hunting Bison“
.
PLOS ONE
.
9
(11): e112884.
Bibcode
:
2014PLoSO...9k2884M
.
doi
:
10.1371/journal.pone.0112884
.
PMC
4229308
.
PMID
25389760
.
- ↑
Power, R. J.; Compion, R. X. Shem (2009).
?Lion predation on elephants in the Savuti, Chobe National Park, Botswana“
.
African Zoology
.
44
(1): 36?44.
doi
:
10.3377/004.044.0104
.
- ↑
Beauchamp 2012
harvnb error: no target: CITEREFBeauchamp2012 (
help
)
- ↑
Dawson, James W. (1988).
?The cooperative breeding system of the Harris' Hawk in Arizona“
. The University of Arizona
. Посетено на
17 November
2017
.
- ↑
Vail, Alexander L.; Manica, Andrea; Bshary, Redouan (23 April 2013). ?Referential gestures in fish collaborative hunting“.
Nature Communications
.
4
(1): 1765.
Bibcode
:
2013NatCo...4.1765V
.
doi
:
10.1038/ncomms2781
.
PMID
23612306
.
- ↑
Yong, Ed (24 April 2013).
?Groupers Use Gestures to Recruit Morays For Hunting Team-Ups“
.
National Geographic
. Посетено на
17 September
2018
.
- ↑
59,0
59,1
Bryce, Caleb M.; Wilmers, Christopher C.; Williams, Terrie M. (2017).
?Energetics and evasion dynamics of large predators and prey: pumas vs. hounds“
.
PeerJ
.
5
: e3701.
doi
:
10.7717/peerj.3701
.
PMC
5563439
.
PMID
28828280
.
- ↑
Majer, Marija; Holm, Christina; Lubin, Yael; Bilde, Trine (2018).
?Cooperative foraging expands dietary niche but does not offset intra-group competition for resources in social spiders“
.
Scientific Reports
.
8
(1): 11828.
Bibcode
:
2018NatSR...811828M
.
doi
:
10.1038/s41598-018-30199-x
.
PMC
6081395
.
PMID
30087391
.
- ↑
?Ambush Predators“
.
Sibley Nature Center
(англиски). Архивирано од
изворникот
на 2021-08-02
. Посетено на
17 September
2018
.
- ↑
Elbroch, L. Mark; Quigley, Howard (10 July 2016).
?Social interactions in a solitary carnivore“
.
Current Zoology
.
63
(4): 357?362.
doi
:
10.1093/cz/zow080
.
PMC
5804185
.
PMID
29491995
.
- ↑
Quenqua, Douglas (11 October 2017).
?Solitary Pumas Turn Out to Be Mountain Lions Who Lunch“
.
The New York Times
(англиски)
. Посетено на
17 September
2018
.
- ↑
Flores, Dan (2016).
Coyote America : a natural and supernatural history
. Basic Books.
ISBN
978-0465052998
.
- ↑
Stow, Adam; Nyqvist, Marina J.; Gozlan, Rodolphe E.; Cucherousset, Julien; Britton, J. Robert (2012).
?Behavioural Syndrome in a Solitary Predator Is Independent of Body Size and Growth Rate“
.
PLOS ONE
.
7
(2): e31619.
Bibcode
:
2012PLoSO...731619N
.
doi
:
10.1371/journal.pone.0031619
.
PMC
3282768
.
PMID
22363687
.
- ↑
?How do Spiders Hunt?“
. American Museum of Natural History. 25 August 2014
. Посетено на
5 September
2018
.
- ↑
?Zooplankton“
. MarineBio Conservation Society
. Посетено на
5 September
2018
.
- ↑
Bar-Yam.
?Predator-Prey Relationships“
. New England Complex Systems Institute
. Посетено на
7 September
2018
.
- ↑
69,0
69,1
69,2
?Predator & Prey: Adaptations“
(PDF)
. Royal Saskatchewan Museum. 2012. Архивирано од
изворникот
(PDF)
на 3 April 2018
. Посетено на
19 April
2018
.
- ↑
Vermeij, Geerat J. (1993).
Evolution and Escalation: An Ecological History of Life
. Princeton University Press. стр. 11 and passim.
ISBN
978-0-691-00080-0
.
- ↑
Getz, W. M. (2011).
?Biomass transformation webs provide a unified approach to consumer-resource modelling“
.
Ecology Letters
.
14
(2): 113?24.
doi
:
10.1111/j.1461-0248.2010.01566.x
.
PMC
3032891
.
PMID
21199247
.
- ↑
Sidorovich, Vadim (2011).
Analysis of vertebrate predator-prey community: Studies within the European Forest zone in terrains with transitional mixed forest in Belarus
. Tesey. стр. 426.
ISBN
978-985-463-456-2
.
- ↑
Angelici, Francesco M. (2015).
Problematic Wildlife: A Cross-Disciplinary Approach
. Springer. стр. 160.
ISBN
978-3-319-22246-2
.
- ↑
Hayward, M. W.; Henschel, P.; O'Brien, J.; Hofmeyr, M.; Balme, G.; Kerley, G.I.H. (2006).
?Prey preferences of the leopard (
Panthera pardus
)“
(PDF)
.
Journal of Zoology
.
270
(2): 298?313.
doi
:
10.1111/j.1469-7998.2006.00139.x
. Архивирано од
изворникот
(PDF)
на 2020-02-29
. Посетено на
2022-04-26
.
- ↑
Pulliam, H. Ronald (1974). ?On the Theory of Optimal Diets“.
The American Naturalist
.
108
(959): 59?74.
doi
:
10.1086/282885
.
- ↑
Sih, Andrew; Christensen, Bent (2001). ?Optimal diet theory: when does it work, and when and why does it fail?“.
Animal Behaviour
.
61
(2): 379?390.
doi
:
10.1006/anbe.2000.1592
.
- ↑
Sprules, W. Gary (1972). ?Effects of Size-Selective Predation and Food Competition on High Altitude Zooplankton Communities“.
Ecology
.
53
(3): 375?386.
doi
:
10.2307/1934223
.
JSTOR
1934223
.
- ↑
Owen-Smith, Norman; Mills, M. G. L. (2008).
?Predator-prey size relationships in an African large-mammal food web“
(PDF)
.
Journal of Animal Ecology
.
77
(1): 173?183.
doi
:
10.1111/j.1365-2656.2007.01314.x
.
PMID
18177336
.
- ↑
Cott 1940
harvnb error: no target: CITEREFCott1940 (
help
)
- ↑
Ruppert, Edward E.; Fox, Richard, S.; Barnes, Robert D. (2004).
Invertebrate Zoology, 7th edition
. Cengage Learning. стр. 153?154.
ISBN
978-81-315-0104-7
.
- ↑
Cetaruk, Edward W. (2005).
?Rattlesnakes and Other Crotalids“
. Во Brent, Jeffrey (уред.).
Critical care toxicology: diagnosis and management of the critically poisoned patient
. Elsevier Health Sciences. стр. 1075.
ISBN
978-0-8151-4387-1
.
- ↑
Barceloux, Donald G. (2008).
Medical Toxicology of Natural Substances: Foods, Fungi, Medicinal Herbs, Plants, and Venomous Animals
. Wiley. стр. 1028.
ISBN
978-0-470-33557-4
.
- ↑
Li, Min; Fry, B.G.; Kini, R. Manjunatha (2005). ?Eggs-Only Diet: Its Implications for the Toxin Profile Changes and Ecology of the Marbled Sea Snake (Aipysurus eydouxii)“.
Journal of Molecular Evolution
.
60
(1): 81?89.
Bibcode
:
2005JMolE..60...81L
.
doi
:
10.1007/s00239-004-0138-0
.
PMID
15696370
.
- ↑
Castello, M. E., A. Rodriguez-Cattaneo, P. A. Aguilera, L. Iribarne, A. C. Pereira, and A. A. Caputi (2009). ?Waveform generation in the weakly electric fish Gymnotus coropinae (Hoedeman): the electric organ and the electric organ discharge“.
Journal of Experimental Biology
.
212
(9): 1351?1364.
doi
:
10.1242/jeb.022566
.
PMID
19376956
.
CS1-одржува?е: пове?е ими?а: список на автори (
link
)
- ↑
Feulner, P. G., M. Plath, J. Engelmann, F. Kirschbaum, R. Tiedemann (2009).
?Electrifying love: electric fish use species-specific discharge for mate recognition“
.
Biology Letters
.
5
(2): 225?228.
doi
:
10.1098/rsbl.2008.0566
.
PMC
2665802
.
PMID
19033131
.
CS1-одржува?е: пове?е ими?а: список на автори (
link
)
- ↑
Catania, Kenneth C. (2015).
?Electric eels use high-voltage to track fast-moving prey“
.
Nature Communications
.
6
(1): 8638.
Bibcode
:
2015NatCo...6.8638C
.
doi
:
10.1038/ncomms9638
.
PMC
4667699
.
PMID
26485580
.
- ↑
Kramer, Bernd (1996).
?Electroreception and communication in fishes“
(PDF)
.
Progress in Zoology
.
42
.
- ↑
Karasov, William H.; Diamond, Jared M. (1988). ?Interplay between Physiology and Ecology in Digestion“.
BioScience
.
38
(9): 602?611.
doi
:
10.2307/1310825
.
JSTOR
1310825
.
- ↑
89,0
89,1
89,2
Jacobs & Bastian 2017
harvnb error: no target: CITEREFJacobsBastian2017 (
help
)
- ↑
Barbosa, Pedro; Castellanos, Ignacio (2005).
Ecology of predator-prey interactions
. Oxford University Press. стр.
78
.
ISBN
9780199874545
.
- ↑
Janis, C. M.; Wilhelm, P. B. (1993). ?Were there mammalian pursuit predators in the Tertiary? Dances with wolf avatars“.
Journal of Mammalian Evolution
.
1
(2): 103?125.
doi
:
10.1007/bf01041590
.
- ↑
92,0
92,1
92,2
Dawkins, Richard;
Krebs, J. R.
(1979). ?Arms races between and within species“.
Proceedings of the Royal Society B: Biological Sciences
.
205
(1161): 489?511.
Bibcode
:
1979RSPSB.205..489D
.
doi
:
10.1098/rspb.1979.0081
.
PMID
42057
.
- ↑
93,0
93,1
Abrams, Peter A. (November 1986). ?Adaptive responses of predators to prey and prey to predators: The failure of the arms-race analogy“.
Evolution
.
40
(6): 1229?1247.
doi
:
10.1111/j.1558-5646.1986.tb05747.x
.
PMID
28563514
.
- ↑
94,0
94,1
94,2
94,3
94,4
Brodie, Edmund D. (July 1999). ?Predator-Prey Arms Races“.
BioScience
.
49
(7): 557?568.
doi
:
10.2307/1313476
.
JSTOR
1313476
.
- ↑
Vermeij, G J (November 1994). ?The Evolutionary Interaction Among Species: Selection, Escalation, and Coevolution“.
Annual Review of Ecology and Systematics
.
25
(1): 219?236.
doi
:
10.1146/annurev.es.25.110194.001251
.
- ↑
Sheriff, Michael J.; Peacor, Scott D.; Hawlena, Dror; Thaker, Maria; Gaillard, Jean?Michel (2020). ?Non-Consumptive Predator Effects on Prey Population Size: A Dearth of Evidence“.
Journal of Animal Ecology
.
89
(6): 1302?1316.
doi
:
10.1111/1365-2656.13213
.
PMID
32215909
.
- ↑
Preisser, Evan L.; Bolnick, Daniel I.; Benard, Michael F. (2005). ?Scared to Death? The Effects of Intimidation and Consumption in Predator?Prey Interactions“.
Ecology
.
86
(2): 501?509.
doi
:
10.1890/04-0719
.
ISSN
0012-9658
.
- ↑
98,0
98,1
Peckarsky, Barbara L.; Abrams, Peter A.; Bolnick, Daniel I.; Dill, Lawrence M.; Grabowski, Jonathan H.; Luttbeg, Barney; Orrock, John L.; Peacor, Scott D.; Preisser, Evan L. (September 2008).
?Revisiting the classics: considering nonconsumptive effects in textbook examples of predator?prey interactions“
.
Ecology
.
89
(9): 2416?2425.
doi
:
10.1890/07-1131.1
.
PMID
18831163
.
- ↑
Lindeman, Raymond L. (1942). ?The Trophic-Dynamic Aspect of Ecology“.
Ecology
.
23
(4): 399?417.
doi
:
10.2307/1930126
.
JSTOR
1930126
.
- ↑
Ordiz, Andres; Bischof, Richard; Swenson, Jon E. (2013). ?Saving large carnivores, but losing the apex predator?“.
Biological Conservation
.
168
: 128?133.
doi
:
10.1016/j.biocon.2013.09.024
.
- ↑
Pimm, S. L.; Lawton, J. H. (1978). ?On feeding on more than one trophic level“.
Nature
.
275
(5680): 542?544.
Bibcode
:
1978Natur.275..542P
.
doi
:
10.1038/275542a0
.
- ↑
Fedriani, J. M.; Fuller, T. K.; Sauvajot, R. M.; York, E. C. (2000). ?Competition and intraguild predation among three sympatric carnivores“.
Oecologia
.
125
(2): 258?270.
Bibcode
:
2000Oecol.125..258F
.
doi
:
10.1007/s004420000448
.
PMID
24595837
.
- ↑
Bond, W. J. (2012). ?11. Keystone species“. Во Schulze, Ernst-Detlef; Mooney, Harold A. (уред.).
Biodiversity and Ecosystem Function
. Springer. стр. 237.
ISBN
978-3642580017
.
- ↑
Botkin, D.; Keller, E. (2003).
Environmental Science: Earth as a living planet
. John Wiley & Sons. стр. 2.
ISBN
978-0-471-38914-9
.
- ↑
105,0
105,1
Ripple, William J.; Beschta, Robert L. (2004). ?Wolves and the Ecology of Fear: Can Predation Risk Structure Ecosystems?“.
BioScience
.
54
(8): 755.
doi
:
10.1641/0006-3568(2004)054[0755:WATEOF]2.0.CO;2
.
- ↑
Neal, Dick (2004).
Introduction to population biology
. Cambridge University Press. стр. 68?69.
ISBN
9780521532235
.
- ↑
Nelson, Erik H.; Matthews, Christopher E.; Rosenheim, Jay A. (July 2004).
?Predators Reduce Prey Population Growth by Inducing Changes in Prey Behavior“
.
Ecology
.
85
(7): 1853?1858.
doi
:
10.1890/03-3109
.
JSTOR
3450359
.
- ↑
Krebs, Charles J.; Boonstra, Rudy; Boutin, Stan; Sinclair, A.R.E. (2001). ?What Drives the 10-year Cycle of Snowshoe Hares?“.
BioScience
.
51
(1): 25.
doi
:
10.1641/0006-3568(2001)051[0025:WDTYCO]2.0.CO;2
.
- ↑
Krebs, Charley; Myers, Judy (2014-07-12).
?The Snowshoe Hare 10-year Cycle ? A Cautionary Tale“
.
Ecological rants
. University of British Columbia
. Посетено на
2 October
2018
.
- ↑
?Predators and their prey“
.
BBC Bitesize
.
BBC
. Посетено на
7 October
2015
.
- ↑
Goel, Narendra S.; Maitra, S. C.; Montroll, E. W. (1971).
On the Volterra and Other Non-Linear Models of Interacting Populations
. Academic Press.
ISBN
978-0122874505
.
- ↑
112,0
112,1
Levin, Simon A.; Carpenter, Stephen R.; Godfray, H. Charles J.; Kinzig, Ann P.; Loreau, Michel; Losos, Jonathan B.; Walker, Brian; Wilcove, David S. (2009).
The Princeton guide to ecology
. Princeton University Press. стр.
204
?209.
ISBN
9781400833023
.
- ↑
Murdoch, William W.; Briggs, Cheryl J.; Nisbet, Roger M. (2013).
Consumer-resource dynamics
. Princeton University Press. стр. 39.
ISBN
9781400847259
.
- ↑
Nowak, Martin; May, Robert M. (2000).
Virus Dynamics : Mathematical Principles of Immunology and Virology
. Oxford University Press. стр. 8.
ISBN
9780191588518
.
- ↑
Genovart, M.; Negre, N.; Tavecchia, G.; Bistuer, A.; Parpal, L.; Oro, D. (2010).
?The young, the weak and the sick: evidence of natural selection by predation“
.
PLOS ONE
.
5
(3): e9774.
Bibcode
:
2010PLoSO...5.9774G
.
doi
:
10.1371/journal.pone.0009774
.
PMC
2841644
.
PMID
20333305
.
- ↑
116,0
116,1
116,2
116,3
Rockwood 2009
harvnb error: no target: CITEREFRockwood2009 (
help
)
- ↑
117,0
117,1
Cushing, J. M. (2005).
?Book Reviews | Mathematics in population biology, by Horst R. Thiene“
(PDF)
.
Bulletin of the American Mathematical Society
.
42
(4): 501?505.
doi
:
10.1090/S0273-0979-05-01055-4
.
- ↑
Thieme, Horst R. (2003).
Mathematics in Population Biology
. Princeton University Press.
ISBN
978-0-691-09291-1
.
- ↑
Kozlov, Vladimir; Vakulenko, Sergey (3 July 2013). ?On chaos in Lotka?Volterra systems: an analytical approach“.
Nonlinearity
.
26
(8): 2299?2314.
Bibcode
:
2013Nonli..26.2299K
.
doi
:
10.1088/0951-7715/26/8/2299
.
- ↑
Sih, Andrew (1987). ?Prey refuges and predator-prey stability“.
Theoretical Population Biology
.
31
: 1?12.
doi
:
10.1016/0040-5809(87)90019-0
.
- ↑
McNair, James N (1986). ?The effects of refuges on predator-prey interactions: A reconsideration“.
Theoretical Population Biology
.
29
(1): 38?63.
doi
:
10.1016/0040-5809(86)90004-3
.
PMID
3961711
.
- ↑
Berryman, Alan A.; Hawkins, Bradford A.; Hawkins, Bradford A. (2006). ?The refuge as an integrating concept in ecology and evolution“.
Oikos
.
115
(1): 192?196.
doi
:
10.1111/j.0030-1299.2006.15188.x
.
- ↑
Cressman, Ross; Garay, Jozsef (2009). ?A predator?prey refuge system: Evolutionary stability in ecological systems“.
Theoretical Population Biology
.
76
(4): 248?57.
doi
:
10.1016/j.tpb.2009.08.005
.
PMID
19751753
.
- ↑
Abrams, P. A. (2000). ?The evolution of predator-prey interactions: theory and evidence“.
Annual Review of Ecology and Systematics
.
31
: 79?105.
doi
:
10.1146/annurev.ecolsys.31.1.79
.
- ↑
125,0
125,1
Grimaldi, David
;
Engel, Michael S.
(2005).
Evolution of the Insects
. Cambridge University Press. стр.
155
?160.
ISBN
978-0-521-82149-0
.
- ↑
Grant, S. W. F.; Knoll, A. H.; Germs, G. J. B. (1991). ?Probable Calcified Metaphytes in the Latest Proterozoic Nama Group, Namibia: Origin, Diagenesis, and Implications“.
Journal of Paleontology
.
65
(1): 1?18.
doi
:
10.1017/S002233600002014X
.
JSTOR
1305691
.
PMID
11538648
.
- ↑
Awramik, S. M. (19 November 1971). ?Precambrian columnar stromatolite diversity: Reflection of metazoan appearance“.
Science
.
174
(4011): 825?827.
Bibcode
:
1971Sci...174..825A
.
doi
:
10.1126/science.174.4011.825
.
PMID
17759393
.
- ↑
Stanley, Steven M. (2008). ?Predation defeats competition on the seafloor“.
Paleobiology
.
34
(1): 1?21.
doi
:
10.1666/07026.1
.
- ↑
Loron, Corentin C.; Rainbird, Robert H.; Turner, Elizabeth C.; Wilder Greenman, J.; Javaux, Emmanuelle J. (2018). ?Implications of selective predation on the macroevolution of eukaryotes: Evidence from Arctic Canada“.
Emerging Topics in Life Sciences
.
2
(2): 247?255.
doi
:
10.1042/ETLS20170153
.
PMID
32412621
.
- ↑
Kelley, Patricia (2003).
Predator--Prey Interactions in the Fossil Record
. Springer. стр. 113?139, 141?176 and passim.
ISBN
978-1-4615-0161-9
.
OCLC
840283264
.
- ↑
Daley, Allison C. (2013). ?Anomalocaridids“.
Current Biology
.
23
(19): R860?R861.
doi
:
10.1016/j.cub.2013.07.008
.
PMID
24112975
.
- ↑
Anderson, P. S. L.; Westneat, M. (2009). ?A biomechanical model of feeding kinematics for Dunkleosteus terrelli (Arthrodira, Placodermi)“.
Paleobiology
.
35
(2): 251?269.
doi
:
10.1666/08011.1
.
- ↑
Carr, Robert K. (2010).
?Paleoecology of Dunkleosteus terrelli (Placodermi: Arthrodira)“
.
Kirtlandia
.
57
.
- ↑
Switeck, Brian (13 April 2012).
?When Tyrannosaurus Chomped Sauropods“
.
Journal of Vertebrate Paleontology
.
25
(2): 469?472.
doi
:
10.1671/0272-4634(2005)025[0469:TRFTUC]2.0.CO;2
. Архивирано од
изворникот
на 24 August 2013
. Посетено на
24 August
2013
.
- ↑
Darimont, C. T.; Fox, C. H.; Bryan, H. M.; Reimchen, T. E. (2015-08-20). ?The unique ecology of human predators“.
Science
.
349
(6250): 858?860.
Bibcode
:
2015Sci...349..858D
.
doi
:
10.1126/science.aac4249
.
PMID
26293961
.
- ↑
Gabriel, Otto; von Brandt, Andres (2005).
Fish catching methods of the world
. Blackwell.
ISBN
978-0-85238-280-6
.
- ↑
Griffin, Emma (2008).
Blood Sport: Hunting in Britain Since 1066
. Yale University Press.
ISBN
978-0300145458
.
- ↑
King, Richard J. (1 October 2013).
The Devil's Cormorant: A Natural History
. University of New Hampshire Press. стр. 9.
ISBN
978-1-61168-225-0
.
- ↑
Glasier, Phillip (1998).
Falconry and Hawking
. Batsford.
ISBN
978-0713484076
.
- ↑
Aegerter, James; Fouracre, David; Smith, Graham C. (2017). Olsson, I Anna S (уред.).
?A first estimate of the structure and density of the populations of pet cats and dogs across Great Britain“
.
PLOS ONE
.
12
(4): e0174709.
Bibcode
:
2017PLoSO..1274709A
.
doi
:
10.1371/journal.pone.0174709
.
PMC
5389805
.
PMID
28403172
.
- ↑
The Humane Society of the United States.
?U.S. Pet Ownership Statistics“
. Архивирано од
изворникот
на 2012-04-07
. Посетено на
27 April
2012
.
- ↑
Liebenberg, Louis (2008). ?The relevance of persistence hunting to human evolution“.
Journal of Human Evolution
.
55
(6): 1156?1159.
doi
:
10.1016/j.jhevol.2008.07.004
.
PMID
18760825
.
- ↑
?Food For Thought“
(PDF)
.
The Life of Mammals
.
British Broadcasting Corporation
. 31 October 2002.
- ↑
Flint, Maria Louise; Dreistadt, Steve H. (1998). Clark, Jack K. (уред.).
Natural Enemies Handbook: The Illustrated Guide to Biological Pest Control
. University of California Press.
ISBN
978-0-520-21801-7
.
- ↑
Johnston, Keith M. (2013).
Science Fiction Film: A Critical Introduction
.
Berg Publishers
. стр. 98.
ISBN
9780857850560
.
- ↑
Schatz, Thomas. ?The New Hollywood“.
Movie Blockbusters
. стр.
25
.
In:
Stringer, Julian (2003).
Movie Blockbusters
.
Routledge
. стр. 15?44.
ISBN
978-0-415-25608-7
.
- ↑
Davison, Peter (1 December 2002).
?Predators and Prey | Selected Poems, 1957-1994 by Ted Hughes“
.
The New York Times
. Посетено на
5 October
2018
.
Hughes's earliest books contained a bewildering profusion of poems between their covers: ... fish and fowl, beasts of the field and forest, vigorous embodiments of predators and prey. Hughes as a student had taken up anthropology, not literature, and he chose to meditate his way into trancelike states of preconsciousness before committing poems to paper. His poems, early or late, enter into the relations of living creatures; they move in close to animal consciousness:
The Thought-Fox,
Esther's Tomcat,
Pike.
- ↑
Gould, Stephen Jay
(1995).
The Tooth and Claw Centennial
.
Dinosaur in a Haystack
. Harmony Books. стр. 63?75.
ISBN
978-0517703939
.
- ↑
149,0
149,1
149,2
149,3
149,4
149,5
Wallner, Astrid (18 July 2005).
?The role of predators in Mythology“
. WaldWissen Information for Forest Management. Архивирано од
изворникот
на 5 October 2018
. Посетено на
5 October
2018
.
translated from Wallner, A. (1998)
Die Bedeutung der Raubtiere in der Mythologie: Ergebnisse einer Literaturstudie
. - Inf.bl. Forsch.bereiches Landsch.okol. 39: 4-5.
- ↑
Kellert, Stephen R.; Black, Matthew; Rush, Colleen Reid; Bath, Alistair J. (1996). ?Human Culture and Large Carnivore Conservation in North America“.
Conservation Biology
.
10
(4): 977?990.
doi
:
10.1046/j.1523-1739.1996.10040977.x
.