Class of enzymes
DNA ligase
is a type of enzyme that facilitates the joining of
DNA
strands together by catalyzing the formation of a
phosphodiester bond
. It plays a role in repairing single-strand breaks in duplex
DNA
in living organisms, but some forms (such as
DNA ligase IV
) may specifically repair double-strand breaks (i.e. a break in both
complementary
strands of DNA). Single-strand breaks are repaired by DNA ligase using the complementary strand of the double helix as a template,
[1]
with DNA ligase creating the final phosphodiester bond to fully repair the DNA.
DNA ligase is used in both
DNA repair
and
DNA replication
(see
Mammalian ligases
). In addition, DNA ligase has extensive use in
molecular biology
laboratories for
recombinant DNA
experiments (see
Research applications
). Purified DNA ligase is used in gene cloning to join DNA molecules together to form
recombinant DNA
.
Enzymatic mechanism
[
edit
]
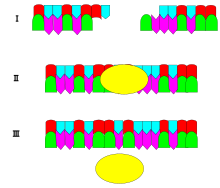 The image demonstrates how ligase (yellow oval) catalyzes two DNA fragment strands. The ligase joins the two fragments of DNA to form a longer strand of DNA by "pasting" them together.
The image demonstrates how ligase (yellow oval) catalyzes two DNA fragment strands. The ligase joins the two fragments of DNA to form a longer strand of DNA by "pasting" them together.
The mechanism of DNA ligase is to form two
covalent
phosphodiester bonds
between
3' hydroxyl ends
of one
nucleotide
("acceptor"), with the
5' phosphate end
of another ("donor"). Two ATP molecules are consumed for each phosphodiester bond formed.
[
citation needed
]
AMP is required for the ligase reaction, which proceeds in four steps:
- Reorganization of activity site such as nicks in DNA segments or Okazaki fragments etc.
- Adenylylation (addition of AMP) of a
lysine
residue in the active center of the enzyme,
pyrophosphate
is released;
- Transfer of the AMP to the 5' phosphate of the so-called donor, formation of a pyrophosphate bond;
- Formation of a phosphodiester bond between the 5' phosphate of the donor and the 3' hydroxyl of the acceptor.
[2]
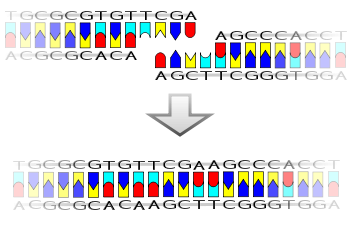 A pictorial example of how a ligase works (with
sticky ends
)
A pictorial example of how a ligase works (with
sticky ends
)
Ligase will also work with
blunt ends
, although higher enzyme concentrations and different reaction conditions are required.
Types
[
edit
]
E. coli
[
edit
]
The
E. coli
DNA ligase is encoded by the
lig
gene. DNA ligase in
E. coli
, as well as most prokaryotes, uses energy gained by cleaving
nicotinamide adenine dinucleotide
(NAD) to create the phosphodiester bond.
[3]
It does not ligate blunt-ended DNA except under conditions of molecular crowding with
polyethylene glycol
, and cannot join RNA to DNA efficiently.
[
citation needed
]
The activity of E. coli DNA ligase can be enhanced by
DNA polymerase
at the right concentrations. Enhancement only works when the concentrations of the DNA polymerase 1 are much lower than the DNA fragments to be ligated. When the concentrations of Pol I DNA polymerases are higher, it has an adverse effect on E. coli DNA ligase
[4]
The DNA ligase from
bacteriophage T4
(a
bacteriophage
that infects
Escherichia coli
bacteria). The T4 ligase is the most-commonly used in laboratory research.
[5]
It can ligate either
cohesive or blunt ends
of DNA, oligonucleotides, as well as RNA and RNA-DNA hybrids, but not single-stranded nucleic acids. It can also ligate
blunt-ended DNA
with much greater efficiency than
E. coli
DNA ligase. Unlike
E. coli
DNA ligase, T4 DNA ligase cannot utilize NAD and it has an absolute requirement for ATP as a cofactor. Some engineering has been done to improve the
in vitro
activity of T4 DNA ligase; one successful approach, for example, tested T4 DNA ligase fused to several alternative DNA binding proteins and found that the constructs with either p50 or
NF-kB
as fusion partners were over 160% more active in blunt-end ligations for cloning purposes than wild type T4 DNA ligase.
[6]
A typical reaction for inserting a fragment into a plasmid vector would use about 0.01 (sticky ends) to 1 (blunt ends) units of ligase. The optimal incubation temperature for T4 DNA ligase is 16 °C.
[
citation needed
]
Bacteriophage T4
ligase
mutants
have increased sensitivity to both
UV
irradiation
[7]
[8]
and the alkylating agent
methyl methanesulfonate
[9]
indicating that DNA ligase is employed in the
repair
of the
DNA damages
caused by these agents.
Mammalian
[
edit
]
In mammals, there are four specific types of ligase.
- DNA ligase 1
: ligates the nascent DNA of the
lagging strand
after the
Ribonuclease H
has removed the RNA primer from the
Okazaki fragments
.
- DNA ligase 3
:
complexes
with
DNA repair
protein
XRCC1
to aid in sealing DNA during the process of
nucleotide excision repair
and recombinant fragments. Of the all known mammalian DNA ligases, only ligase 3 has been found to be present in mitochondria.
- DNA ligase 4
: complexes with
XRCC4
. It catalyzes the final step in the
non-homologous end joining
DNA double-strand break repair pathway. It is also required for
V(D)J recombination
, the process that generates diversity in
immunoglobulin
and
T-cell receptor
loci during
immune system
development.
- DNA ligase 2: A purification artifact resulting from proteolytic degradation of DNA ligase 3. Initially, it has been recognized as another DNA ligase and it is the reason for the unusual nomenclature of DNA ligases.
[10]
DNA ligase from
eukaryotes
and some microbes uses
adenosine triphosphate
(ATP) rather than NAD.
[3]
Thermostable
[
edit
]
Derived from a thermophilic bacterium, the enzyme is stable and active at much higher temperatures than conventional DNA ligases. Its half-life is 48 hours at 65 °C and greater than 1 hour at 95 °C. Ampligase DNA Ligase has been shown to be active for at least 500 thermal cycles (94 °C/80 °C) or 16 hours of cycling.
10
This exceptional thermostability permits extremely high hybridization stringency and ligation specificity.
[11]
Measurement of activity
[
edit
]
There are at least three different units used to measure the activity of DNA ligase:
[12]
- Weiss unit
- the amount of ligase that catalyzes the exchange of 1 nmole of
32
P from inorganic
pyrophosphate
to ATP in 20 minutes at 37
°
C. This is the one most commonly used.
- Modrich-Lehman unit
- this is rarely used, and one unit is defined as the amount of enzyme required to convert 100 nmoles of d(A-T)
n
to an exonuclease-III resistant form in 30 minutes under standard conditions.
- Many commercial suppliers of ligases use an arbitrary unit based on the ability of ligase to ligate cohesive ends. These units are often more subjective than quantitative and lack precision.
Research applications
[
edit
]
DNA ligases have become indispensable tools in modern molecular biology research for generating
recombinant DNA
sequences. For example, DNA ligases are used with
restriction enzymes
to insert DNA fragments, often
genes
, into
plasmids
.
Controlling the optimal temperature is a vital aspect of performing efficient recombination experiments involving the ligation of cohesive-ended fragments. Most experiments use T4 DNA Ligase (isolated from
bacteriophage T4
), which is most active at 37 °C.
[13]
However, for optimal ligation efficiency with cohesive-ended fragments ("sticky ends"), the optimal enzyme temperature needs to be balanced with the
melting temperature
T
m
of the sticky ends being ligated,
[14]
the
homologous pairing
of the sticky ends will not be stable because the high temperature disrupts
hydrogen bonding
. A ligation reaction is most efficient when the sticky ends are already stably annealed, and disruption of the annealing ends would therefore result in low ligation efficiency. The shorter the
overhang
, the lower the T
m
.
Since blunt-ended DNA fragments have no cohesive ends to anneal, the melting temperature is not a factor to consider within the normal temperature range of the ligation reaction. The limiting factor in blunt end ligation is not the activity of the ligase but rather the number of alignments between DNA fragment ends that occur. The most efficient ligation temperature for blunt-ended DNA would therefore be the temperature at which the greatest number of alignments can occur. The majority of blunt-ended ligations are carried out at 14-25 °C overnight. The absence of stably annealed ends also means that the ligation efficiency is lowered, requiring a higher ligase concentration to be used.
[14]
A novel use of DNA ligase can be seen in the field of nano chemistry, specifically in DNA origami. DNA based self-assembly principles have proven useful for organizing nanoscale objects, such as biomolecules, nanomachines, nanoelectronic and photonic component. Assembly of such nano structure requires the creation of an intricate mesh of DNA molecules. Although DNA self-assembly is possible without any outside help using different substrates such as provision of catatonic surface of Aluminium foil, DNA ligase can provide the enzymatic assistance that is required to make DNA lattice structure from DNA over hangs.
[15]
History
[
edit
]
The first DNA ligase was purified and characterized in 1967 by the Gellert, Lehman, Richardson, and Hurwitz laboratories.
[16]
It was first purified and characterized by Weiss and Richardson using a six-step chromatographic-fractionation process beginning with elimination of cell debris and addition of streptomycin, followed by several Diethylaminoethyl (DEAE)-cellulose column washes and a final phosphocellulose fractionation. The final extract contained 10% of the activity initially recorded in the
E. coli
media; along the process it was discovered that ATP and Mg++ were necessary to optimize the reaction. The common commercially available DNA ligases were originally discovered in
bacteriophage T4
,
E. coli
and other
bacteria
.
[17]
Disorders
[
edit
]
Genetic deficiencies in human DNA ligases have been associated with clinical syndromes marked by immunodeficiency, radiation sensitivity, and developmental abnormalities,
[16]
LIG4 syndrome
(Ligase IV syndrome) is a rare disease associated with mutations in DNA ligase 4 and interferes with dsDNA break-repair mechanisms. Ligase IV syndrome causes immunodeficiency in individuals and is commonly associated with microcephaly and marrow hypoplasia.
[18]
A list of prevalent diseases caused by lack of or malfunctioning of DNA ligase is as follows.
Xeroderma pigmentosum
[
edit
]
Xeroderma pigmentosum
, which is commonly known as XP, is an inherited condition characterized by an extreme sensitivity to ultraviolet (UV) rays from sunlight. This condition mostly affects the eyes and areas of skin exposed to the sun. Some affected individuals also have problems involving the nervous system.
[19]
Ataxia-telangiectasia
[
edit
]
Mutations in the ATM gene cause
ataxia?telangiectasia
. The ATM gene provides instructions for making a protein that helps control cell division and is involved in DNA repair. This protein plays an important role in the normal development and activity of several body systems, including the nervous system and immune system. The ATM protein assists cells in recognizing damaged or broken DNA strands and coordinates DNA repair by activating enzymes that fix the broken strands. Efficient repair of damaged DNA strands helps maintain the stability of the cell's genetic information. Affected children typically develop difficulty walking, problems with balance and hand coordination, involuntary jerking movements (chorea), muscle twitches (myoclonus), and disturbances in nerve function (neuropathy). The movement problems typically cause people to require wheelchair assistance by adolescence. People with this disorder also have slurred speech and trouble moving their eyes to look side-to-side (oculomotor apraxia).
[20]
Fanconi Anemia
[
edit
]
Fanconi anemia
(FA) is a rare, inherited blood disorder that leads to bone marrow failure. FA prevents bone marrow from making enough new blood cells for the body to work normally. FA also can cause the bone marrow to make many faulty blood cells. This can lead to serious health problems, such as
leukemia
.
[21]
Bloom syndrome
[
edit
]
Bloom syndrome
results in skin that is sensitive to sun exposure, and usually the development of a butterfly-shaped patch of reddened skin across the nose and cheeks. A skin rash can also appear on other areas that are typically exposed to the sun, such as the back of the hands and the forearms. Small clusters of enlarged blood vessels (telangiectases) often appear in the rash; telangiectases can also occur in the eyes. Other skin features include patches of skin that are lighter or darker than the surrounding areas (hypopigmentation or hyperpigmentation respectively). These patches appear on areas of the skin that are not exposed to the sun, and their development is not related to the rashes.
As a drug target
[
edit
]
In recent studies, human DNA ligase I was used in
Computer-aided drug design
to identify DNA ligase inhibitors as possible therapeutic agents to treat cancer.
[22]
Since excessive cell growth is a hallmark of cancer development, targeted chemotherapy that disrupts the functioning of DNA ligase can impede adjuvant cancer forms. Furthermore, it has been shown that DNA ligases can be broadly divided into two categories, namely, ATP- and NAD
+
-dependent. Previous research has shown that although NAD
+
-dependent DNA ligases have been discovered in sporadic cellular or viral niches outside the bacterial domain of life, there is no instance in which a NAD
+
-dependent ligase is present in a
eukaryotic
organism. The presence solely in non-eukaryotic organisms, unique substrate specificity, and distinctive domain structure of NAD+ dependent compared with ATP-dependent human DNA ligases together make NAD
+
-dependent ligases ideal targets for the development of new antibacterial drugs.
[16]
See also
[
edit
]
References
[
edit
]
- ^
Pascal JM, O'Brien PJ, Tomkinson AE, Ellenberger T (November 2004). "Human DNA ligase I completely encircles and partially unwinds nicked DNA".
Nature
.
432
(7016): 473?8.
Bibcode
:
2004Natur.432..473P
.
doi
:
10.1038/nature03082
.
PMID
15565146
.
S2CID
3105417
.
- ^
Lehman IR (November 1974). "DNA ligase: structure, mechanism, and function".
Science
.
186
(4166): 790?7.
Bibcode
:
1974Sci...186..790L
.
doi
:
10.1126/science.186.4166.790
.
PMID
4377758
.
S2CID
86549159
.
- ^
a
b
Foster JB, Slonczewski J (2010).
Microbiology: An Evolving Science
(Second ed.). New York: W. W. Norton & Company.
ISBN
978-0-393-93447-2
.
- ^
Yang Y, LiCata VJ (February 2018).
"Pol I DNA polymerases stimulate DNA end-joining by Escherichia coli DNA ligase"
.
Biochemical and Biophysical Research Communications
.
497
(1): 13?18.
doi
:
10.1016/j.bbrc.2018.01.165
.
PMID
29409896
.
- ^
"Ligases"
(PDF)
.
Enzyme Resources Guide
. Promega Corporation. pp. 8?14.
- ^
Wilson RH, Morton SK, Deiderick H, Gerth ML, Paul HA, Gerber I, Patel A, Ellington AD, Hunicke-Smith SP, Patrick WM (July 2013).
"Engineered DNA ligases with improved activities in vitro"
.
Protein Engineering, Design & Selection
.
26
(7): 471?8.
doi
:
10.1093/protein/gzt024
.
PMID
23754529
.
- ^
Baldy MW (1968). "Repair and recombination in phage T4. II. Genes affecting UV sensitivity".
Cold Spring Harbor Symposia on Quantitative Biology
.
33
: 333?8.
doi
:
10.1101/sqb.1968.033.01.038
.
PMID
4891973
.
- ^
Baldy MW (February 1970). "The UV sensitivity of some early-function temperature-sensitive mutants of phage T4".
Virology
.
40
(2): 272?87.
doi
:
10.1016/0042-6822(70)90403-4
.
PMID
4909413
.
- ^
Baldy MW, Strom B, Bernstein H (March 1971).
"Repair of alkylated bacteriophage T4 deoxyribonucleic acid by a mechanism involving polynucleotide ligase"
.
Journal of Virology
.
7
(3): 407?8.
doi
:
10.1128/JVI.7.3.407-408.1971
.
PMC
356131
.
PMID
4927528
.
- ^
Tomkinson, Alan E; Sallmyr, Annahita (5 September 2013).
"Structure and function of the DNA ligases encoded by the mammalian LIG3 gene"
.
Gene
.
531
(2): 150?157.
doi
:
10.1016/j.gene.2013.08.061
.
PMC
3881560
.
PMID
24013086
.
- ^
"Ampligase- Thermostable DNA Ligase"
.
www.epibio.com
. Archived from
the original
on 2017-06-19
. Retrieved
2017-05-15
.
- ^
Russell DW, Sambrook J (2001). "Chapter 1: Plasmids and Their Usefulness in Molecular Cloning".
Molecular cloning: a laboratory manual
. Vol. 1 (3rd ed.). Cold Spring Harbor, N.Y: Cold Spring Harbor Laboratory. pp. 1?159.
ISBN
978-0-87969-577-4
.
- ^
Baneyx F, Lucotte G (1993).
Introduction to Molecular Cloning Techniques
. Chichester: John Wiley & Sons. p. 156.
ISBN
978-0-471-18849-0
.
- ^
a
b
Tabor S (May 2001). "DNA ligases".
Current Protocols in Molecular Biology
. Chapter 3: Unit 3.14.
doi
:
10.1002/0471142727.mb0314s08
.
ISBN
978-0-471-14272-0
.
PMID
18265223
.
S2CID
23944826
.
- ^
Bhanjadeo MM, Nayak AK, Subudhi U (2017). "Surface-assisted DNA self-assembly: An enzyme-free strategy towards formation of branched DNA lattice".
Biochemical and Biophysical Research Communications
.
485
(2): 492?498.
doi
:
10.1016/j.bbrc.2017.02.024
.
PMID
28189681
.
- ^
a
b
c
Shuman S (June 2009).
"DNA ligases: progress and prospects"
.
The Journal of Biological Chemistry
.
284
(26): 17365?9.
doi
:
10.1074/jbc.R900017200
.
PMC
2719376
.
PMID
19329793
.
- ^
Weiss B, Richardson CC (April 1967).
"Enzymatic breakage and joining of deoxyribonucleic acid, I. Repair of single-strand breaks in DNA by an enzyme system from Escherichia coli infected with T4 bacteriophage"
.
Proceedings of the National Academy of Sciences of the United States of America
.
57
(4): 1021?8.
Bibcode
:
1967PNAS...57.1021W
.
doi
:
10.1073/pnas.57.4.1021
.
PMC
224649
.
PMID
5340583
.
- ^
Altmann T, Gennery AR (October 2016).
"DNA ligase IV syndrome; a review"
.
Orphanet Journal of Rare Diseases
.
11
(1): 137.
doi
:
10.1186/s13023-016-0520-1
.
PMC
5055698
.
PMID
27717373
.
- ^
"xeroderma pigmentosum"
.
Genetics Home Reference
. Retrieved
2017-05-15
.
- ^
"ataxia-telangiectasia"
.
Genetics Home Reference
. Retrieved
2017-05-15
.
- ^
"What Is Fanconi Anemia?"
. NHLBI, NIH
. Retrieved
2017-05-15
.
- ^
Tomkinson AE, Howes TR, Wiest NE (June 2013).
"DNA ligases as therapeutic targets"
.
Translational Cancer Research
.
2
(3).
PMC
3819426
.
PMID
24224145
.
External links
[
edit
]
|
|---|
| 6.5
: Phosphoric Ester
| |
|---|
| 6.6
: Nitrogen-Metal
| |
|---|
|
|---|
| Activity
| |
|---|
| Regulation
| |
|---|
| Classification
| |
|---|
| Kinetics
| |
|---|
| Types
| |
|---|