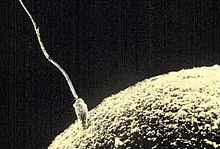
 Spermium und Eizelle
Spermium und Eizelle
Ein
Spermium
(
Plural
Spermien
),
Spermatozoon
(Plural
Spermatozoen
) oder
Spermatozoid
(Plural
Spermatozoiden
), auch
Samenfaden
oder
Samenzelle
genannt, ist eine Form von
Gameten
(Keimzellen), namlich eine zu eigenstandiger Bewegung fahige, reife Keimzelle, die bei der Vereinigung mit einer unbeweglichen, meist großeren Keimzelle (
Eizelle
) zu deren
Befruchtung
fuhrt. Mannliche Keimzellen, die nicht zu eigenstandiger Bewegung fahig sind, werden als
Spermatien
(Singular
Spermatium
) bezeichnet.
Bei Menschen werden Spermien von einem mannlichen Individuum meistens in großer Zahl in den Samenkanalchen der Hoden produziert und sind wesentlich kleiner als die zu befruchtende Eizelle der Frau, weil sie im Gegensatz zur Eizelle keine großeren
Plasmamengen
und
dotterhaltigen
Nahrstoffe
enthalten. Zu unterscheiden sind die Begriffe
Spermium
und
Sperma
. Das
Sperma
(die Flussigkeit) besteht aus dem
Seminalplasma
mit den darin enthaltenen Spermien und etlichen
Epithelzellen
der
Hodenkanalchen
.
Verschiedene Zelltypen, so auch die Spermatozyten, besitzen eine Form des
Immunprivilegs
.
[1]
Dieses Privileg verhindert eine
Autoimmunreaktion
gegen die Spermatozyten und ist zur Vorbeugung von Infektionen ausschlaggebend. Vergleiche mit Mausen haben gezeigt, dass besonders Menschen bei einer Storung dieses Privilegs haufig an einer
Orchitis
zu erkranken scheinen, welche wiederum eine mogliche Ursache fur
Unfruchtbarkeit
ist.
Trotz der historisch bedingt synonymen Bezeichnungsverwendung ist das Spermium
nicht
homolog zum
Samen
der Pflanzen, der bereits den pflanzlichen Embryo enthalt, sondern es handelt sich um einen Gameten, welchem bei Pflanzen die generative
Zelle
in einem
Pollenkorn
funktionell entspricht.
Als Zelle hatte
Adolph von La Valette-St. George
das Spermium (ebenso wie das
Ei
) 1865/1866 nachgewiesen.
[2]
 Feinstruktur eines humanen Spermatozoons
Feinstruktur eines humanen Spermatozoons
Menschliche Spermien konnen mit einem gewohnlichen Lichtmikroskop bereits bei einer 100-, besser aber 400-fachen Vergroßerung ohne Einfarbung beobachtet werden. Beim menschlichen Spermium handelt es sich um eine begeißelte Zelle, die einen Kopfteil mit
haploidem
Chromosomensatz in einem
Zellkern
(entweder mit X- oder Y-Chromosom), ein Mittelstuck (?Hals“ mit
Zentrosom
und darumliegenden
Mitochondrien
-Paketen) sowie eine
Geißel
(auch als ?Schwanz“ bezeichnet) besitzt. An der der Geißel gegenuberliegenden Seite, also beim Schwimmen an der Vorderseite des Spermiums, liegt der Kopf, der fur das Eindringen in die
Eizelle
zustandig ist. Am außersten Ende des Spermienkopfs befindet sich die Kopfkappe (
Akrosom
), die mit
Enzymen
gefullt ist, die das Durchdringen der Ei-Membran erleichtern. Mittel- und Schwanzstuck enthalten den Achsenfaden (
axial filament
). Dieses Filamentbundel entspringt am Zentriol und besteht wie eine Geißel aus zwei Zentral- und neun peripheren Doppelfilamenten.
Die Große der Spermien variiert bei den einzelnen Arten stark. Wahrend die Riesenspermien von
Ostrakoden
(Muschelkrebse) 7 mm lang sind und damit bis zu zehnmal so lang wie die Muschelkrebse selbst werden konnen,
[3]
besitzen menschliche Spermien nur eine Lange von etwa 60 μm: der Kopfteil ist hierbei etwa 5 μm lang und 3 μm breit, wahrend die Geißel einschließlich ?Hals“ rund 50 μm lang ist. Die Große der Spermien kann sogar innerhalb der gleichen
Art
variieren, je nachdem, ob ein Mannchen mehrere Nebenbuhler hat oder nicht. So hat man bei
Froschen
festgestellt, dass die Spermiengroße und damit auch die Lange der Geißel zunimmt, wenn das betreffende Mannchen sich mit anderen Mannchen um die Befruchtung der Eier eines Froschweibchens auseinanderzusetzen hat. Die Spermien mit der langsten Geißel, die am schnellsten schwimmen konnen, haben dabei die großte Chance, als erste die vom Weibchen ins Wasser abgegebenen Froscheier zu erreichen.
Nicht nur bei
Sußwasser-Muschelkrebsen
, sondern auch bei einigen anderen Arten im Tierreich gibt es Riesenspermien. Diese sind teilweise um ein Vielfaches langer als ihr Produzent. Sie kommen bei einigen Arten von Wurmern, Schmetterlingen und Wasserwanzen vor. Den Großenrekord halt dabei die
Taufliege
Drosophila bifurca
. Ihre Samenzellen messen mit bis zu 58 Millimetern Lange mehr als das Zehnfache der Korperlange des Insekts.
[4]
Bei niederen
Krebsen
und etlichen
Spinnentieren
konnen die Spermien insgesamt kugelformig sein. Der
Spulwurm
besitzt nagelformige Spermien mit einem Glanzkorper aus spezifischen
Eiweißen
. Andere
Rundwurmer
und auch
Milben
haben amoboid bewegliche Spermien. Bei den
Zehnfußkrebsen
(
Decapoda
) kommt eine Art ?Explosionseinrichtung“ in Form eines Sprungfedermechanismus vor, der das Spermium in die Eizelle katapultiert.
Bei
Wirbeltieren
werden Spermien im
Epithel
der
Hodenkanalchen
des
Hodens
produziert. Siehe
Spermatogenese
.
 Phasenweise Darstellung des Eindringens des Spermiums in eine Eizelle
Phasenweise Darstellung des Eindringens des Spermiums in eine Eizelle
Bei
Galenos
(2. Jahrhundert n. Chr.) herrschte die Vorstellung von in den Hoden gebildetem mannlichen und in den ?weiblichen Hoden“, den Ovarien, gebildetem weiblichen ?Sperma“
[5]
bzw. Samen.
[6]
Spermatozoen wurden mikroskopisch vom Medizinstudenten
Johan Ham
(† nach 1723
[7]
) entdeckt. Er kam zu
Antoni van Leeuwenhoek
, der weiterfuhrende Untersuchungen machte und 1677 einen Brief mit der Entdeckung an die
Royal Society
schickte.
[8]
[9]
Leeuwenhoek benannte die neu entdeckten ?Partikel“ wie schon zuvor von ihm entdeckte
Bakterien
als
Animalcula
, also (Samen-)Tierchen, und ordnete sie den
Infusorien
zu, er erkannte bereits richtig, dass sie im Hoden gebildet werden mussen. Seiner Vorstellung nach als einer der Begrunder der
Praformationslehre
handelte es sich aber um etwas wie Menschen-Larven. Der gesamte Mensch ware demnach im Spermium als ?
Homunculus
“ bereits vorgebildet und musse nur noch heranwachsen, er bestritt jede Bedeutung der Mutter und der mutterlichen Eier (die
Eizelle
wurde erst 1827 entdeckt); sein Konkurrent
Jan Swammerdam
vertrat jedoch die gegenteilige Ansicht. In der Forschung der folgenden Jahrhunderte tobte ein erbitterter Streit zwischen den ?Ovisten“, die den Keim des kunftigen Menschen im Ei, und den ?Animalculisten“, die ihn im Spermium lokalisierten. Beide glaubten an eine Praformation des Menschen in dem jeweiligen Keim, moglicherweise sogar immer kleiner eingeschachtelt, und so auf die ersten Menschen Adam oder Eva zuruckgehend.
[10]
Den Ausdruck
Spermatozoon
(griechisch fur Samentier, Samenlebewesen) verwendete erstmals 1826 der Embryologe
Karl Ernst von Baer
, der die fertile Funktion dieser ?Lebewesen im Sperma“ damals zu Recht fur unbewiesen hielt.
[11]
1842 veroffentlichte der Schweizer Biologe
Albert von Kolliker
seine Untersuchungen an Spermien in dem Werk
Untersuchungen uber die Bedeutung der Samenfaden
.
Dass Spermien in das Ovum eindringen, wurde mikroskopisch erstmals 1843 von
Martin Barry
beobachtet und berichtet.
[12]
Das Spermium des Mannes besteht aus
- einem Kopfteil, der den
haploiden
Chromosomensatz und zwischen 2682 und 2886 verschiedene
mRNA
-Molekule im
Zellkern
enthalt,
- einem Mittelstuck mit vier bis funf
Mitochondrien
, die die
Energie
in Form von
ATP
-Molekulen fur die Fortbewegung liefern,
- einem beweglichen Schwanzteil mit langsverlaufendem
Fibrillensystem
aus
Mikrotubuli
zur Fortbewegung.
Menschliche Spermien dienen ? wie die Spermatozoen der anderen Organismen ? der
Befruchtung
einer weiblichen
Eizelle
. Sie werden nach ihrer Fertigstellung (
Spermatogenese
) zunachst im mannlichen
Nebenhoden
gelagert, um von dort aus uber den
Samenleiter
und die
Harnrohre
bei der
Ejakulation
wahrend des mannlichen
Orgasmus
ausgestoßen zu werden. Rund 300 Millionen von ihnen landen in der weiblichen
Scheide
. Von der Scheide aus gelangt nur ein kleiner Teil der Spermien, die zur Befruchtung vorgesehen sind, uber den
Eileiter
zur Eizelle, dem weitaus großeren Teil gelingt der hindernisreiche Weg bis in diese Region nicht, oder aber er ist fur andere Funktionen vorgesehen (vergl.
Spermienkonkurrenz
).
Auf dem Weg zur Eizelle werden die Spermien wahrscheinlich
chemotaktisch
von
Progesteron
oder duftahnlichen Substanzen, dem
pH-Wert
und Temperaturunterschieden geleitet.
[13]
Aufgenommen werden die Reize einerseits von Molekulen der großen Familie der G-Protein-gekoppelten Rezeptoren in der
Membran
des Anfangsteils des Spermienschwanzes, die z. T. mit denen in den
Riechzellen
unserer
Nase
identisch sind (Geruchsrezeptoren), andererseits durch den CatSper-Ionenkanal, der auch durch viele niedermolekulare Substanzen aktiviert werden kann.
[14]
Experimentell konnte gezeigt werden: Bindet der Duftstoff Bourgeonal
[15]
(Maiglockchenduft) an den
OR1D2
, steigt im Innern des Spermiums die
Calcium
-Konzentration. Gleiches gilt nach aktuellen Erkenntnissen fur eine Bindung und Aktivierung des CatSper-Kanals.
[14]
Dies hat zur Folge, dass das Spermium seine Schwimmrichtung andert und gleichzeitig die Schwimmgeschwindigkeit verdoppelt.
[15]
Dabei sind insbesondere die Veranderungen der Calcium-Konzentrationen, nicht deren absolute Hohe, fur die Richtung der Fortbewegung verantwortlich.
[16]
[17]
Es ist jedoch hochst unwahrscheinlich, dass der naturliche Bindungspartner des Bourgeonal-Rezeptors der einzige ?Wegweiser“ zur Eizelle ist; eher ist von mehreren Faktoren auszugehen.
[13]
Wegen der vielen Hindernisse erreichen unter Normalbedingungen nur etwa 300 Spermien diejenige Stelle am Ende des Eileiters, an der die Eizelle auf ihre Befruchtung wartet. Die Eizelle lasst sich allerdings nur von einem einzigen Spermium befruchten. Bei der
Befruchtung
dringt der Inhalt des Spermienkopfes in die Eizelle ein, die dadurch
diploid
wird und nun
Zygote
heißt. Nach neueren Erkenntnissen beeinflussen die zusammen mit dem Zellkern des Spermiums in die Eizelle eingedrungenen mannlichen
mRNA
-Molekule die Entwicklung des aus der Zygote entstehenden
Embryos
. 2018 wurde entdeckt, dass in seltenen Fallen auch Mitochondrien des Spermiums in die Eizelle gelangen konnen.
Menschliche Spermien unter einem
Lichtmikroskop
 Darstellung der Krafte, die bei der rhythmischen Fortbewegung eines Spermiums wirken.
Darstellung der Krafte, die bei der rhythmischen Fortbewegung eines Spermiums wirken.
Die menschlichen Spermien besitzen eine aktiv bewegliche
Geißel
. Im 17. Jahrhundert beschrieb der niederlandische Naturforscher
Antoni van Leeuwenhoek
als erster menschliche Samenzellen und erkannte mittels
Lichtmikroskop
auch die langliche Geißel, mit deren Hilfe sie sich fortbewegen. Nach Aussage dieses Forschers peitschen die Spermien beim Schwimmen ihre Geißeln schlangenahnlich wie
Aale
im Wasser. Deshalb galt uber 300 Jahre in den Lehrbuchern, dass die beiden auf der Geißel markierten Punkte sich nach oben (linker Punkt) und unten (rechter Punkt) bewegen. Diese Geschwindigkeit wird aufgeteilt in den Teil, der parallel (V[par]) zum Abschnitt der Geißel verlauft, und den Teil, der senkrecht (V[senkr]) dazu verlauft. Die daraus resultierende Kraft F wird wiederum in zwei Teile aufgeteilt, namlich den Teil der Kraft, der parallel zur Bewegungsrichtung des Spermiums wirkt (F[horiz]), und den Teil, der vertikal zur Bewegungsrichtung wirkt (F[vert]). Die Summe der beiden Krafte, die parallel zur Bewegungsrichtung des Spermiums gerichtet sind, bilden die vorantreibende Kraft.
[18]
Laut einer neuen Studie drehen sich jedoch bei der Fortbewegung die Geißeln eher wie ein
Korkenzieher
immer in eine Richtung.
[19]
Das Forscherteam um
Hermes Gadelha
hat hochauflosende 3-D-Aufnahmen von Spermien gemacht und konnte dabei die tatsachliche Bewegungsart beobachten. Demnach ist die Geißel schief und schlangelt nur auf eine Seite. Damit sich die Spermien auf diese Weise nicht nur im Kreis herum bewegen und somit den Weg zur Eizelle verfehlen, drehen sie sich als Ganzes wie Korkenzieher oder
Kreisel
, gleichen damit die einseitige Schlagbewegung der Geißel aus, und bewegen sich vorwarts. Mit dem von van Leeuwenhoek und auch heute oft verwendeten normalen Mikroskop war und ist das nicht zu erkennen, denn die schnellen und synchronisierten Drehbewegungen sehen dabei so aus, als bewege sich die Geißel symmetrisch von Seite zu Seite. Die neuen Erkenntnisse durch Beobachtungen mit einem 3-D-Mikroskop konnen nach Ansicht der Forscher auch praktische Folgen haben. Da die Ursache von mehr als der Halfte aller Falle von Unfruchtbarkeit beim Mann liegt, ist es sehr bedeutend und grundlegend, genau zu verstehen, wie der Schwanz von Spermien sich tatsachlich bewegt und funktioniert, um in Zukunft Exemplare mit Fehlfunktion erkennen zu konnen.
[19]
Nach einer Reifungsdauer von etwa 10 Wochen von der Spermatogonie zur Spermatozoe
[20]
(ausgereiftes Spermium) kann ein Spermium bis zu einem Monat im Spermadepot des Mannes uberdauern. An der Luft konnen Spermien je nach Umweltbedingungen (Licht, Temperatur, Feuchtigkeit) bis zu 24 Stunden uberleben. Sobald das
Ejakulat
mit den Spermien trocknet, sterben diese ab. Es kann also keine Befruchtung mittels eingetrocknetem Sperma stattfinden.
[21]
Dies liegt daran, dass die Spermien zum Uberleben die Flussigkeit benotigen, die sie umgibt: das Seminalplasma. Solange darin die Bewegungsfahigkeit der Samenzellen gesichert ist, sind sie lebensfahig.
[22]
Durch den Ausgleich der pH-Werte von Vagina (pH 4?5) und Zervixschleim und Sperma (pH 6?8) ist es den Spermien moglich, in dem an sich ≫feindlichen≪ Milieu zu uberleben. Unter optimalen Bedingungen in den Buchten der Zervixschleimhaut konnen Spermien dort bis zu sieben Tage uberleben. Von den durchschnittlich 250 Millionen Spermien pro Samenerguss erreichen nur ca. 500 bis 800 die Eileiter. Ein Grund hierfur ist der Zervixschleim, durch den die nicht schwimmfahigen Spermien gefiltert werden.
[23]
Eine Abweichung vom optimalen, leicht basischen
pH-Wert
(7,2?7,8)
[24]
fuhrt zum Absterben der Spermien. Die meisten in mechanischen und chemischen
Empfangnisverhutungsmethoden
verwendeten
Spermizide
arbeiten auf dieser Basis. Die im Genitalbereich haufig verwendeten pH-neutralen Pflegeprodukte sind nicht spermizid und fur eine empfangnisverhutende Nachsorge ungeeignet.
Die Spermienqualitat in den
Industriestaaten
geht weltweit seit Jahren zuruck. Bei 60 % der untersuchten Schweizer zwischen 18 und 22 Jahren liegt mindestens ein Kennwert unterhalb des Grenzwerts der
Weltgesundheitsorganisation
(WHO) fur gute Zeugungsfahigkeit.
[25]
Kritiker, darunter Dolores Lamb, die stellvertretende Direktorin der urologischen Klinik der
Cornell University
, wenden ein, dass die Qualitat der verwendeten Daten der meisten Studien zur Spermienqualitat unzureichend sei und sich die verwendete Zahlweise uber die Jahre stark geandert habe. Dadurch seien die meisten Studien zusammen mit darauf basierenden Metastudien sinnlos. Als Gegenbeispiel wird die unter sehr kontrollierten Bedingungen ausgefuhrte Studie von Elisabeth Carlsen in Danemark angefuhrt, bei der zwischen 1996 und 2010 sogar eine leichte Steigerung der Spermienkonzentration der Probanden festgestellt wurde.
[26]
Die zweigeißeligen Gameten der mannlichen Geschlechtsorganen (Antheridien) von
Moosen
,
Farnen
und
Barlapppflanzen
werden Spermatozoide genannt. In der Regel gelangen diese durch Regen auf das
Archegonium
, das weibliche Geschlechtsorgan der Sporenpflanzen. Hier konnen sie sich bis zu mehreren Millimetern fortbewegen, bis sie die
Zygote
erreichen.
Hornmoose
besitzen rechtsdrehende Spermatozoide. Nach der Befruchtung bildet sich der
Sporophyt
.
Der Begriff
Spermienkonkurrenz
beschreibt die
Konkurrenz
von Spermien eines oder mehrerer Mannchen um die Chance zur Befruchtung einer
Eizelle
. Spermienkonkurrenz entsteht, weil die Mannchen aller Tier
arten
sehr viel mehr Spermien produzieren als die Weibchen ihrer Art
befruchtungsfahige
Eizellen.
Eine weitere Variante, die zuerst bei Insekten beobachtet wurde und auch bei den Saugetieren vorkommen soll, besteht darin, dass die Mannchen nicht nur befruchtungsfahige Spermien ejakulieren, sondern auch bewegungsunfahige und solche, die durch Zelloberflachenmolekule konkurrierende Spermien abtoten. Die bewegungsunfahigen Spermien blockieren den Weg etwaig vorhandener Fremdspermien, so dass sie schlechter an den weiblichen Schleimhautoberflachen entlang zur Eizelle wandern konnen. Sogenannte ?Killerspermien“ suchen gezielt nach Fremdspermien, deren Oberflachenstruktur nicht der eigenen entspricht, und toten diese chemisch ab.
[27]
[28]
Eine defensive Strategie besteht darin, ein Weibchen moglichst oft zu begatten. Dies fuhrt dazu, dass sich im Korper des Weibchens zu jeder Zeit eine großere Menge eigener Spermien befinden. So haben die Spermien eines Konkurrenten eine geringere Wahrscheinlichkeit, zur Befruchtung zu gelangen.
Spermien
und
Sperma
werden oft als
Samen
bezeichnet. Dies kann in die Irre fuhren, denn ein
Same
ist ein (oft in Fruchtfleisch eingebettetes) Verbreitungsorgan der hoheren Pflanzen, das aus einem ruhenden pflanzlichen
Embryo
besteht, der von
Nahrgewebe
und einer
Samenschale
umgeben ist.
Die Verwendung der Bezeichnung
Same
oder
Samen
fur die Spermien leitet sich aus der
Bibel
ab, wobei sie dort nicht den Anspruch erhebt, wissenschaftlich korrekt zu sein, sondern eher verwandtschaftliche Abstammung betonen will, und dies auf eine fur damalige Verhaltnisse verstandliche Weise. Das hebraische Wort fur
Same
(
???
zera
) wird dort unterschiedslos fur Pflanzen, Tiere und den Menschen gebraucht. So empfangt einerseits die Frau den mannlichen Samen (Num 5, 28) oder erweckt ihn beim erotischen Spiel (Gen 19, 32 und 34), andererseits wird das Land mit den Samen der Feldfruchte besat (Dtn 29, 22; Ez 36, 9).
Zudem beschrankte sich in antiken und mittelalterlichen Vorstellungen das Vorhandensein eines ?Samens“ als Keimzelle nicht nur auf das mannliche Geschlecht.
[29]
Das
Corpus Hippocraticum
sah den Zeugungsstoff als Samen an, welcher von beiden Geschlechtern sowohl in weiblicher als auch mannlicher Form beigesteuert wird. Das Geschlecht des dabei gezeugten Kindes hinge vom Verhaltnis des starkeren zum schwacheren Samen ab.
[30]
Aus dem alten
Agypten
stammt die falsche Vorstellung, dass der mannliche
Same
bereits der Mensch
in nuce
sei, der im Mutterleib quasi wie in einer Nahrlosung nur noch heranzureifen braucht. Diese Vorstellung wurde in Antike und Neuzeit als
Praformationslehre
vertreten.
Schließlich bezeichnet das Wort
Same
auch die Nachkommenschaft selbst. Wenn die Bibel vom Samen
Abrahams
spricht, dann sind damit die aus Abraham hervorgegangenen Nachkommen gemeint (Jes 41, 8; Jer 33, 26). All diese Bedeutungen sind hier nicht gemeint. Neuere Schulbucher sprechen nicht mehr vom
Samenleiter
, sondern ausdrucklich vom
Spermienleiter
.
Tatsachlich bedeutet bereits das griechische Wort σπ?ρμα (Sperma) nichts anderes als ?Samen“.
[31]
Das findet sich auch in botanischen Bezeichnungen wie
Angiospermen
fur ?Bedecktsamer“ wieder. Die Mehrdeutigkeit besteht also in mehreren Sprachen.
- Spermatophore
(Samenpakete) dienen bei manchen Tierarten der Spermienubertragung.
- Ein
Spermiogramm
wird bei Menschen durch Analyse des
Ejakulats
erstellt und dient der Beurteilung der Zeugungsfahigkeit des Mannes.
- Britta Hahnel:
ReproTier-Kompetenzverbund prasentiert: Die kleine Spermienfibel: Großenmessung an Spermien verschiedener Tierarten
. durchgefuhrt und dokumentiert im Institut fur Fortpflanzung landwirtschaftlicher Nutztiere Schonow e. V., Mensch & Buch, Hahnel 2007,
ISBN 978-3-86664-186-0
.
- Robie H. Harris:
Was jetzt kommt ist … einfach irre! Ein Buch uber Eier und Spermien, Geburt, Babys und Zusammenleben
. Beltz & Gelberg, Weinheim 2002.
ISBN 3-407-75319-5
.
- Vivien Marx:
Das Samenbuch, alles uber Spermien, Sex und Fruchtbarkeit
(=
Fischer-Taschenbuch.
Band 14140). Fischer, Frankfurt am Main 1999,
ISBN 3-596-14140-0
.
- 1938:
Walther Schonfeld
:
Um die Entdeckung der menschlichen Samenfaden
(Ludwig von Hammen aus Danzig ? Johan Ham aus Arnheim [Holland] ? Antony van Leeuwenhoek aus Delft). In:
Archiv fur Dermatologie und Syphilis
. Band 178, Nummer 3, 1938, S. 358?372,
ISSN
0365-6020
.
- 1950: Olaf W. Dietz:
Die Zahl der Spermien im Ejakulat des Ziegenbockes in der Abhangigkeit zur Sexualpause
. Dissertation Universitat Leipzig, Veterinar-medizinische Fakultat, 15. Dezember 1950,
DNB
481838465
.
- 1971: Charles A. Joel:
Historical survey of research on spermatozoa from antiquity to the present.
In:
Fertility disturbances in men and women.
Basel 1971, S. 3?43.
- 1974: Stephan Schulte-Wrede:
Raster-Elektronenmikroskopie von Spermien des Hausschafs ?Ovis ammon aries L.“
Dissertation Universitat Munchen 1974,
DNB
780784022
(Aus:
Zeitschrift fur Zellforschung und mikroskopische Anatomie.
Nr. 134, 1972,
ISSN
0340-0336
, S. 105?127, zusammen mit Rudolf Wetzstein).
- 1983: Ulrich Wirth:
Spermien und Spermatogenese bei Nematoden und die Bedeutung der Spermien fur die Phylogenetik der Metazoen
. Dissertation, Universitat Freiburg, Freiburg im Breisgau 1983,
DNB
840435703
.
- 1990: Heike Rauhaus:
Untersuchungen zur Morphologie und Lebend-Tot-Farbung von Spermien einiger Haustierarten
. Dissertation, Universitat Munchen, 1990,
DNB
901541443
.
- 1995: Dirk Schulze Bertelsbeck:
Die Bedeutung von Spermienantikorpern in Serum und auf Spermien fur die Diagnose der immunologisch bedingten Infertilitat
. Dissertation Universitat Munster (Westfalen) 1995,
DNB
946147442
.
- 1996: Manuela Quandt:
Inhibition und Stimulation der Spermienmigration im in vitro Spermien-Mukus-Interaktionsmodell.
Dissertation, Universitat Heidelberg, 1996,
DNB
949085073
.
- 1996: Stefan Hans Uhlich:
Vergleich von Spermien nach Praparation mit Glaswollfiltration oder Percoll-Dichtegradientenzentrifugation: eine elektronenmikroskopische Untersuchung
. Dissertation, Universitat Ulm, 1996,
DNB
949658227
.
- 1999: World Health Organization (Hrsg.):
WHO-Laborhandbuch zur Untersuchung des menschlichen Ejakulates und der Spermien-Zervikalschleim-Interaktion
(Originaltitel:
WHO Laboratory Manual for the Examination of Human Semen and Sperm Cervical Mucus Interaction.
ubersetzt von Eberhard Nieschlag und Susan Nieschlag in Zusammenarbeit mit Monika Bals-Pratsch). Springer, Berlin/ Heidelberg/ New York u. a. 1999,
ISBN 3-540-66335-5
.
- 2001: Andrea Wagner:
Das funktionelle Spermienreservoir im Saugetier. Charakterisierung der kohlenhydratvermittelten Vorgange der Spermien-Oviduktbindung beim Schwein
. Dissertation, Tierarztliche Hochschule Hannover, 2001,
DNB
964080087
(
online
PDF, kostenfrei, 113 Seiten, 1,3 MB).
- 2002: Brigitte Reimesch:
Untersuchungen zum Einfluss von Coenzym Q10 und einer Mischung aus Coenzym Q10 und Vitamin C, in vitro, auf die Beweglichkeit der Spermien.
(
Mikrofiche
). Dissertation, Universitat Erlangen-Nurnberg, 2002,
DNB
964741318
.
- 2002: Steffen Klaus Meurer:
Molekularbiologische und immunologische Charakterisierung von Chemorezeptoren in Saugetier-Spermien
(=
Berichte des Forschungszentrums Julich
). Forschungszentrum Julich, Zentralbibliothek, Julich 2002
ISSN
0944-2952
(= Dissertation, Universitat Koln, 2002).
- 2003: Johannes Solzin:
Chemotaxis von Seeigel-Spermien, kinetische Messungen intrazellularer Botenstoffe
(=
Forschungszentrum Julich: Berichte des Forschungszentrums Julich.
Band 4030). Dissertation, Universitat Koln, 2003,
DNB
968795285
.
- 2010: Britta Verena Behr:
The biotechnological potential for manipulating offspring sex in the rhinoceros and the elephant.
Freie Universitat, Berlin 2010,
ISBN 978-3-86664-702-2
(Dissertation, FU Berlin, 2009, Jornalnummer 3291
online
PDF, kostenfrei, 126 Seiten, 3,8 MB (englisch)).
- ↑
S. Zhao, W. Zhu, S. Xue, D. Han:
Testicular defense systems: immune privilege and innate immunity.
In:
Cellular & molecular immunology.
Band 11, Nr. 5, September 2014, S. 428?437,
doi
:
10.1038/cmi.2014.38
,
PMID 24954222
,
PMC 4197207
(freier Volltext).
- ↑
Paul Diepgen
,
Heinz Goerke
:
Aschoff
/Diepgen/Goerke: Kurze Ubersichtstabelle zur Geschichte der Medizin.
7., neubearbeitete Auflage. Springer, Berlin/Gottingen/Heidelberg 1960, S. 41.
- ↑
Siehe
Erfolgsmodell Riesenspermium.
und
Wettlauf der Giganten ? Riesenspermien in Mikrofossilien nachgewiesen
.
- ↑
Spektrum der Wissenschaft:
Das uralte Erbe der Riesenspermien.
September 2009, S. 14?16.
- ↑
Wolfgang Gerlach:
Das Problem des weiblichen Samens in der antiken und mittelalterlichen Medizin.
In:
Sudhoffs Archiv.
Band 30, Heft 4?5, 1938, S. 177?193.
- ↑
Erna Lesky
:
Galen als Vorlaufer der Hormonforschung.
In:
Centaurus.
Band 1, 1950/51, S. 156?162, hier: S. 159?161.
- ↑
Paul Diepgen
,
Heinz Goerke
:
Aschoff
: Kurze Ubersichtstabellen zur Geschichte der Medizin.
7., neubearbeitete Auflage. Springer, Berlin/ Gottingen/ Heidelberg 1960, S. 24.
- ↑
W. Schonfeld:
Um die Entdeckung der menschlichen Samenfaden (Ludwig von Hammen aus Danzig ? Johan Ham aus Arnheim [Holland] ? Antony van Leeuwenhoek aus Delft)
. In:
Archives of Dermatological Research.
Band 178, Nr. 3, 1938, S. 358?372,
DOI:10.1007/BF02061155
.
- ↑
Observationes D. Anthonii Lewenhoeck, de Natis e semine genitali Animalculis.
In:
Philosophical Transactions of the Royal Society of London.
Band 12, Nr. 1677, S. 1040?1046,
doi
:
10.1098/rstl.1677.0068
(
Volltext
).
- ↑
Hans Fischer:
Die Geschichte der Zeugung- und Entwicklungstheorien im 17. Jahrhundert.
In:
Gesnerus.
Band 2, Nr. 2, 1945, S. 49?80,
doi
:
10.5169/seals-520562
.
- ↑
zitiert aus
Karl Friedrich Burdach
:
Die Physiologie als Erfahrungswissenschaft.
Band 1. Voss, Leipzig 1826, S. 90, (vgl. Th. Schmuck:
Baltische Genesis. Die Grundlegung der modernen Embryologie.
Shaker, Aachen 2009,
ISBN 978-3-83228-781-8
, S. 182).
- ↑
M. Barry:
Spermatozoa Observed within the Mammiferous Ovum.
In:
Philosophical Transactions of the Royal Society of London.
Band 133, Nr. 1843, S. 33?33,
doi
:
10.1098/rstl.1843.0005
.
- ↑
a
b
Michael Eisenbach, Laura C. Giojalas:
Sperm guidance in mammals ? An unpaved road to the egg.
In:
Nature Reviews Molecular Cell Biology
.
Band 7, Nr. 4, Mai 2006, S. 276?285,
DOI: 10.1038/nrm1893
.
- ↑
a
b
Christoph Brenker, Normann Goodwin, Ingo Weyand u. a.:
The CatSper channel: a polymodal chemosensor in human sperm.
In:
The EMBO journal.
(
EMBO J
) Band 31, Nr. 7, 4. April 2012, S. 1654?1665,
doi:10.1038/emboj.2012.30
.
- ↑
a
b
M. Spehr, G. Gisselmann, A. Poplawski,
Hanns Hatt
:
Identification of a testicular odorant receptor mediating human sperm chemotaxis.
In:
Science.
Band 299, Nr. 5615, 28. Marz 2003, S. 2054?2058,
DOI:10.1126/science.1080376
.
- ↑
Luis Alvarez, Luru Dai, Benjamin M. Friedrich, Nachiket D. Kashikar, Ingo Gregor, Rene Pascal,
U. Benjamin Kaupp
:
The rate of change in Ca
2+
concentration controls sperm chemotaxis.
In:
Journal of Cell Biology
Band 196, 2012,
DOI:10.1083/jcb.201106096
.
- ↑
Mitch Leslie:
As the sperm turns.
In:
Journal of Cell Biology
Band 196, Nr. 5, Marz 2012, S. 548?548,
DOI:10.1083/jcb.1965iti3
.
- ↑
Len Fisher:
Reise zum Mittelpunkt des Fruhstuckseis. Streifzuge durch die Physik der alltaglichen Dinge
(=
Bastei-Lubbe-Taschenbuch.
Band 77218). Bastei Lubbe Stars, Bergisch Gladbach 2007,
ISBN 3-404-77218-0
.
- ↑
a
b
Hermes Gadelha, Paul Hernandez-Herrera, Fernando Montoya, Alberto Darszon u. a.:
Human sperm uses asymmetric and anisotropic flagellar controls to regulate swimming symmetry and cell steering.
In:
Science Advances.
Band 6, Nr. 31, 31. Juli 2020, Artikel: eaba5168,
DOI:10.1126/sciadv.aba5168
(
Volltext als PDF
).
- ↑
Renate Lullmann-Rauch:
Histologie: Verstehen, Lernen, Nachschlagen; 10 Tabellen.
Thieme, Stuttgart/ New York 2003,
ISBN 3-131-29241-5
, S. 404.
- ↑
Peter Thiel:
Sperma.
Auf:
maennerberatung.de
vom 5. Juni 2017; abgerufen am 13. August 2020.
- ↑
Svenja Hauke:
Wie lange uberleben Spermien außerhalb des Korpers?
Auf:
gelbeseiten.de/ratgeber
vom 12. Februar 2019; abgerufen am 15. Januar 2023.
- ↑
Sarah Gruber:
Basics Gynakologie und Geburtshilfe
(=
Basics.
) 2., durchgesehene Auflage, Elsevier/ Urban & Fischer, Munchen 2007,
ISBN 3-437-42157-3
, S. 72.
- ↑
Renate Lullmann-Rauch:
Histologie: Verstehen, Lernen, Nachschlagen; 10 Tabellen.
Stuttgart/ New York 2003, S. 416 ff.
- ↑
Katrin Zoefel:
Studie zur Fruchtbarkeit - Besorgnis wegen schlechter Spermienqualitat.
In:
srf.ch
.
22. Mai 2019,
abgerufen am 22. Mai 2019
.
- ↑
Tom Kattwinkel:
Die Spur der verlorenen Spermien.
In:
Zeit Online
.
31. Juli 2023,
abgerufen am 1. August 2023
.
- ↑
G. A. Parker:
Sperm competition and its evolutionary consequences in the insects.
In:
Biological Reviews.
Band 45, Nr. 4, 1970, S. 525?567.
- ↑
Robin Baker
, Mark A. Bellis:
Human Sperm Competition: Copulation, masturbation and infidelity.
Springer, Berlin/ Heidelberg /New York 1994,
ISBN 978-0-4124-5430-1
.
- ↑
Wolfgang Gerlach:
Das Problem des weiblichen Samens in der antiken und mittelalterlichen Medizin.
In:
Sudhoffs Archiv.
Nr. 30, 1938, S. 177?193.
- ↑
Jutta Kollesch
,
Diethard Nickel
:
Antike Heilkunst. Ausgewahlte Texte aus den medizinischen Schriften der Griechen und Romer.
Philipp Reclam junior, Leipzig 1979 (=
Reclams Universal-Bibliothek.
Band 771); 6. Auflage ebenda 1989,
ISBN 3-379-00411-1
, S. 75 f. (
Hippokrates, Uber den Samen, Kap. 6?8.
).
- ↑
Bernhard Kytzler, Lutz Redemund, Nikolaus Eberl:
Unser tagliches Griechisch. Lexikon des griechischen Spracherbes.
3. Auflage, von Zabern, Mainz 2007,
ISBN 978-3-8053-2816-6
.